Sommaire de la page
Structure résumée
Directeur du département: Carlos Bernstein, DR CNRS
19 chercheurs et enseignants chercheurs permanents 4 post-doctorants et 27 doctorants
Écologie du Comportement et Dynamique des Populations
|
|
Anciens membres de l’équipe ayant quitté le laboratoire entre 2002 et 2005:
M. Ney-Nifle, CR CNRS, partie au 31 août 2005 au laboratoire Leibniz – IMAG à Grenoble ; H. Ploye MCU, (retraite au 1er septembre 2004).
Écologie évolutive et biologie des populations d’insectes
|
|
Anciens membres de l’équipe ayant quitté le laboratoire entre 2002 et 2005:
B. Soula (thèse, 2002) enseignant dans un lycée
Écologie évolutive des populations
|
|
Anciens membres de l’équipe ayant quitté le laboratoire entre 2002 et 2005:
N. Bahi (thèse, 2003), en post-doc en Tunisie à l’Institut Pasteur, ; P. Blanchard (thèse 2002) post-doc aux Kerguelen ; S. Devillard (thèse 2004) post-doc à l’ONCFS ; N. Pettorelli (thèse 2002) post-doc au Canada ; N. Valière (thèse 2002) en formation au CAPES ; M. Bourgarel (thèse 2004) CR au CIRAD ; S. Rossi (thèse 2005) Vétérinaire Inspecteur.
Introduction
L’exploitation durable des ressources naturelles, la lutte contre les maladies émergentes, la conservation et gestion de la biodiversité imposent de nouveaux défis à l’écologie en termes de gestion, contrôle et de mise en place de mesures de conservation des populations. Cette gestion ne peut pas être faite sur la base de recettes empiriques ou selon la puissance relative des différents groupes d’intérêt de notre société. Elle doit être basée sur la meilleure connaissance scientifique et les meilleures méthodologies disponibles. Les domaines de l’écologie des populations et des peuplements a donc un rôle fondamental à jouer dans les prochaines décennies. L’étude des aspects fondamentaux de cette discipline n’est pas un passe-temps intellectuel mais un apport indispensable pour une société qui doit faire face à un avenir incertain en termes d’environnement.
L’écologie ne se résume pas à un simple recueil de descriptions phénoménologiques et de recettes empiriques mais constitue une science à part entière reposant sur un corps théorique rigoureux, des méthodologies élaborées et de forts liens avec d’autres disciplines (génétique des populations, mathématiques, biochimie…). Comme on le verra par la suite, toute l’activité de notre département est guidée par cette conception de l’écologie. Nous cherchons donc à participer au développement du cadre théorique de notre discipline et à mettre à l’épreuve les hypothèses qui en découlent. Nous avons aussi établi de forts liens scientifiques avec des disciplines telles que les mathématiques, la statistique, l’analyse moléculaire, la biochimie via des collaborations avec des spécialistes de premier ordre.
L’un des leviers les plus importants du développement de l’écologie a été la prise de conscience que de nombreuses caractéristiques des êtres vivants sont le résultat de l’évolution par la sélection naturelle et que les structures en populations sont la conséquence des interactions entre les individus. Cette approche évolutive de la nature, qui est le noyau central de l’Écologie Évolutive, a été fondamentale pour le développement de toute l’écologie comme discipline scientifique. Les résultats théoriques et expérimentaux ont démontré l’existence d’une interdépendance fondamentale ente les aspects évolutifs et la dynamique des populations et communautés. Une approche évolutive devient ainsi fondamentale pour la compréhension générale du fonctionnement de la nature.
La pensée adaptative guide nos hypothèses et travaux expérimentaux ; elle constitue également le meilleur moyen de détecter les contraintes qui limitent l’univers des possibles adaptatifs. Cette conception de l’écologie est centrale à toute notre activité. Ceci est illustré à travers quatre grands axes :
- Évolution des traits d’histoire de vie, stratégies de reproduction et structuration des populations
- Relations interspécifiques
- Comportement et dynamique des populations
- Gestion et conservation dans le cadre des relations interspécifiques
Ces quatre axes résument l’activité du département indépendamment des groupes taxonomiques et des équipes impliquées. Cette unité d’approche et de problématiques nous à conduit à présenter nos activités plutôt en termes de grands axes scientifiques qu’en termes d’équipes.
Évolution des traits d’histoire de vie, stratégies de reproduction et structuration des populations animales
Un des objectifs de l’Écologie Évolutive est de comprendre les pressions sélectives responsables de la diversité des traits d’histoire de vie mais aussi les interactions entre traits (trade-offs) et les contraintes (e.g., inertie phylogénétique, contraintes allométriques…) qui limitent leur optimisation. Pour démontrer la valeur adaptative d’un trait ou d’un compromis évolutif entre traits, il est nécessaire d’estimer les coûts et les bénéfices qui lui sont associés. Les stratégies de développement et de reproduction illustrent parfaitement ces compromis. L’évolution des traits d’histoire de vie et la dynamique des populations constituent deux composantes d’une même thématique qu’il est important de considérer en interaction. Par exemple, la variabilité des traits d’histoire de vie et les différences génétiques associées influencent les structures et la dynamique des populations.
Ces thèmes ont été abordés au sein de notre département au travers des cinq questions développées ci-dessous.
1) Évolution de la reproduction sexuée
En raison des coûts liés à la reproduction sexuée, son ubiquité au sein des règnes animal et végétal constitue un paradoxe et demeure une question ouverte en biologie évolutive.
Des lignées sexuées versus asexuées adaptées à des conditions locales stables
Chez l’hyménoptère parasitoïde Venturia canescens les modes de reproduction sexuée (arrhénotoquie) et asexuée (thélytoquie) se rencontrent en sympatrie dans le milieu naturel (Schneider et al., 2002, Schneider (thèse 2003), Amat (thèse 2004). L’arrhénotoquie prédomine en conditions naturelles alors que les populations en milieux anthropiques sont exclusivement thélytoques. Nous avons testé si les conditions environnementales locales (i.e. de l’environnement préférentiel) pour chaque mode reproductif avaient sélectionné pour des différences dans les stratégies d’exploitation des hôtes. Pour chaque mode reproductif, les parasitoïdes devraient être plus sensibles aux facteurs qui présentent une forte variabilité dans leur environnement préférentiel. Ainsi, nous avons montré (Fig1) que les femelles sexuées sont plus sensibles à des diminutions de température, s’attardent plus longtemps et exploitent plus intensément le patch d’hôtes où elles se trouvent. Ce résultat est en accord avec le comportement attendu de parasitoïdes en réponse à des signes de détérioration des conditions dans un futur proche. (Amat et al., 2005).
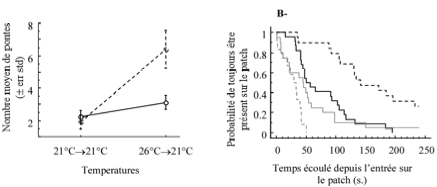
Figure 1 : Le comportement d’exploitation des patchs par des femelles de V. canescens asexuées et sexuées est étudié en fonction des conditions de température auxquelles elles sont soumises : conditionnement (48 heures à une certaine température, avant le test) des femelles et exploitation des patchs (période de test) à 21°C (21°C→21°C) ou conditionnement à 26°C et exploitation des patchs à 21°C (26°C→21°C). A- nombre moyen de ponte (± err std) effectué par les femelles asexuées (trait plein) et sexuées (trait pointillé) en fonction des conditions de température. B- probabilité pour une femelle asexuée (trait plein) ou sexuée (trait pointillé) de toujours être présente sur le patch en fonction du temps écoulé depuis son entrée sur ce patch. En noir, lorsque les femelles subissent une diminution de température avant d’exploiter les patchs (26°C→21°C) et en gris lorsque le conditionnement et l’exploitation sont effectués à la même température (21°C→21°C).
Plasticité « pile ou face » et allocation au sexe : une réponse à l’imprévisibilité environnementale
Une stratégie pour répondre à un degré d’imprévisibilité environnementale élevé réside en une variation phénotypique exprimée aléatoirement par un même génotype à chaque génération (« coin-flipping plasticity », Cooper et Kaplan, 1982). Cependant, il est difficile de distinguer ce type d’adaptation d’une variation non adaptée. En collaboration avec des collègues de Rennes et étrangers, nous venons d’apporter l’indication de l’existence de cette plasticité en ce qui concerne l’allocation au sexe chez des femelles de pucerons Rhopalosiphum padi (Halkett et al., 2004). En automne, les femelles de cette espèce sont confrontées à deux options: soit produire des descendants sexués qui s’accouplent et pondent des œufs diapausants qui supportent les froids hivernaux, soit continuer de produire des femelles parthénogénétiques sensibles au froid. Si l’hiver est doux, la production de ce type de femelles est associée à un bénéfice en termes de taux d’accroissement, mais un hiver froid tuera toute la descendance. À l’aide d’un modèle de programmation dynamique, nous avons montré qu’une femelle, qui produit aléatoirement à la fois des descendants sexués et parthénogénétiques, serait sélectionnée par l’imprévisibilité de l’occurrence des hivers froids. L’investissement dans la reproduction sexuée est proportionnel à la sévérité de l’hiver, et la variance dans le pic de la date de production des individus sexués dépend de l’imprévisibilité climatique. Ces prédictions ont été validées à l’aide de captures d’insectes in natura et de relevés météorologiques dans les sites étudiés.
2) Évolution des systèmes d’appariement
S’il est admis que le système d’appariement est une composante des stratégies bio-démographiques des espèces, les conséquences du système d’appariement sur le fonctionnement des populations sont encore mal évaluées. Les développements de l’outil moléculaire ont permis deux avancées majeures dans l’étude de ses conséquences : (i) une identification précise du système d’appariement des espèces étudiées; (ii) une caractérisation du succès reproducteur réel des individus. Nos travaux ont concerné ces deux thèmes.
À la suite de la mise en évidence de paternité hors couples (EPP : « extra-pair paternity ») chez la marmotte alpine, espèce socialement monogame, nos travaux se sont focalisés sur l’analyse des causes évolutives de ces EPP (thèse d’Aurélie Cohas). Les principaux résultats indiquent que les EPP ne sont pas observés en absence de subordonnés mâles, ce qui suggère que le mâle dominant est alors capable de « contrôler » la femelle et d’empêcher les copulations hors couples (EPC). Lorsque l’environnement social permet les EPC, nous avons montré que les EPP conduisent à une augmentation de diversité génétique des individus et des portées (Cohas et al., 2005).
Chez la population des chats de l’archipel des Kerguelen et utilisant les marqueurs moléculaires pour déterminer la paternité des portées, nous avons montré que le système d’appariement est de type monogame. L’hypothèse pour expliquer ce résultat étonnant chez le chat est qu’il s’agit d’un processus densité-dépendant. Du fait de la très faible densité (de l’ordre d’un chat au km2), un mâle ne pourrait pas monopoliser plus d’une femelle. Cette variation du système d’appariement selon l’habitat (polygyne en milieu rural, promicuiste en milieu urbain) a théoriquement des implications sur la taille efficace des populations en raison de la différence de variance dans le système d’appariement. Il a été possible de le démontrer empiriquement à partir de données recueillies dans les populations de chats rurales et urbaines (Kaeuffer et al. 2004). La taille efficace d’une population polygyne (forte variance du succès reproducteur entre mâles) est bien inférieure à celle d’une population promiscuiste, caractérisée par une variance du succès reproducteur entre mâles plus faible. Ces résultats couplés à d’autres (Devillard et al., 2003a, 2004a, (Devillard, thèse 2004), permettent d’asseoir définitivement l’extrême plasticité biodémographique du chat domestique.
3) Reproduction immédiate ou différée : approches déterministe et stochastique
Selon l’hétérogénéité spatiale et temporelle de leur environnement, les individus doivent « choisir » d’investir dans une reproduction immédiate ou dans une reproduction différée selon les avantages et les coûts associés à chacune des stratégies. Dans certains contextes écologiques, c’est un compromis entre ces deux stratégies, qui a été sélectionné.
En environnement déterministe
Chez les parasitoïdes, ce compromis (trade-off) est en partie déterminé par les décisions de chercher des hôtes (gain immédiat de fitness) ou de la nourriture (gain différé de fitness via une augmentation de la survie). Alors que la recherche d’hôtes est bien documentée, les sources de nourriture, les indices utilisés par les parasitoïdes pour chercher de la nourriture et leurs conséquences comportementales sont peu ou pas connus. Dans un premier temps, à l’aide d’analyses biochimiques d’animaux capturés sur le terrain nous avons montré que V. canescens s’alimentait fréquemment en conditions naturelles (Casas et al., 2003, Fig. 2). Une des sources de nourriture est constituée des fruits qui abritent les hôtes à attaquer (Desouhant et al., 2005). Dans un deuxième temps, des expériences au laboratoire et sur le terrain ont mis en évidence la capacité de cet insecte à détecter des indices chimiques associés à la nourriture et aux hôtes. Les guêpes s’orientent préférentiellement vers les sites qui contiennent à la fois des hôtes et de la nourriture. De cette façon, elles réduisent le temps et l’énergie associés à la recherche de nourriture, ressources qui peuvent être alloués à l’exploitation des hôtes (Desouhant et al., 2005).
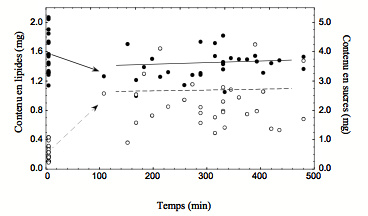
Figure 2 : Changement des contenus en lipides (axe gauche des ordonnées, points noirs, ligne pleine) et en sucres (axe droit des ordonnées, points blancs et ligne pointillée) pour des guêpes Venturia canescens lâchées sur le terrain puis recapturées le même jour.Le groupe contrôle (t=0) correspond à des guêpes congelées juste avant le lâcher. Les droites de régression sont pour les guêpes recapturées (unité en mg/0.02 mm de longueur de tibia). L’origine des flèches correspond à la moyenne des valeurs pour les groupes contrôle. Comparé au contrôle, le niveau moyen des réserves lipidiques des guêpes recapturées diminue alors que celui de sucres augmente. Ceci suggère que les réserves énergétiques en lipides sont fixées à la naissance alors que celles de sucres sont renouvelées via une activité alimentaire fréquente (sur miellat ou nectar).
La diapause prolongée est également un moyen de différer sa reproduction. Les effets de la diapause prolongée sur la dynamique des systèmes hôtes-parasitoïdes ont été étudiés d’un point de vue théorique (Corley et al., 2004) en incorporant des patrons réalistes de développement des hôtes et des parasitoïdes. Contrairement à des résultats antérieurs, nos modèles prédisent que la diapause prolongée se traduit par une stabilité réduite.
Nous verrons dans la section suivante que la prise en compte des stratégies de diapause est également indispensable pour comprendre l’évolution en environnement stochastique.
En environnement stochastique
La plupart des études théoriques sur l’évolution des traits d’histoires de vie supposent un environnement stable et/ou prévisible. Cependant, il est important de confronter les approximations des modèles déterminismes aux prédictions d’approches qui intègrent la composante aléatoire (i.e. imprévisible) de la variabilité environnementale. À l’aide d’une approche multi-disciplinaire associant modélisation, expérimentations et suivi à long terme de populations naturelles, nous avons étudié cette problématique en nous focalisant sur l’évolution des stratégies de diapause et de ponte chez un insecte, le balanin de la châtaigne Curculio elephas.
Nos travaux antérieurs ont démontré l’existence d’une plasticité dans la durée du cycle de vie du balanin de la châtaigne. Chez le balanin, à chaque génération, deux types de larves sont produites par un même génotype: des larves à cycle court (diapause de courte durée) qui émergent en tant qu’adultes après une année, et des larves à cycle long (diapause prolongée) qui produisent des adultes après deux, trois ou quatre ans. Nous avons démontré que le « choix » d’induire un cycle court ou long était corrélé au poids des larves et à la quantité de réserve énergétique (lipides) (Menu et Desouhant, 2002 ; Soula et Menu, 2005). Cette corrélation entre durée du cycle et réserves énergétiques permet aux larves à cycle long de compenser une perte énergétique supplémentaire durant la diapause prolongée (Soula et Menu, 2004). En conséquence, les adultes issues des larves à cycles longs présentent des performances similaires aux adultes issus de larves à cycle court (Soula et Menu, 2003, Fig. 3).
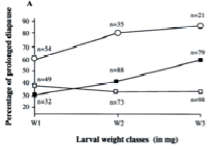
Figure 3 : Probabilité d’entrer en diapause prolongée selon le poids et la date de sortie hors des fruits chez le balanin de la châtaigne Curculio elephas (Coléoptère Curculionidés) (Menu et Desouhant, 2002). Le poids et la date de sortie larvaire hors des fruits sont deux indicateurs des conditions de développement larvaire dans les fruits (nourriture, température…).Les dates de sortie des fruits sont regroupées en trois cohortes : carrés blancs : cohorte 1 (10-21 octobre), carrés noirs : cohorte 2 (22-31 octobre), ronds blancs : cohorte 3 (1-27 novembre). Trois classes de poids :≤80 mg (W1), >80≤100 mg (W2) et >100 mg (W3). n représente le nombre total d’individus pour chaque groupe
Une étude de micro-respirométrie, a permis de démontrer, chez le balanin, que la diapause prolongée n’est pas une simple extension de la diapause hivernale. Durant la première année dans le sol, les « chemins développementaux » des larves à cycle long et des larves à cycle court présentent la même séquence de développement et divergent seulement en juin-juillet, moment où les larves à cycle court se nymphosent et celles à cycle long induisent une deuxième diapause. Cette deuxième diapause dure au moins une année, et doit être considérée comme une diapause prolongée intervenant secondairement dans le cycle de l’insecte. Ce résultat renforce notamment notre hypothèse d’une induction de la diapause prolongée basée sur les ressources lipidiques (Soula et Menu, 2005).
4) Évolution de la sexe ratio
L’évolution de la sex-ratio dans les populations naturelles a depuis longtemps intrigué les biologistes de l’évolution. Au plan théorique, plusieurs mécanismes ont été proposés pour expliquer des biais de sex-ratio au niveau de la population. Ces biais possibles à l’échelle de la population sont la résultante des biais produits au niveau individuel, et Trivers et Willard (1973) ont été les premiers à suggérer que les femelles devraient ajuster la sex-ratio dans leur portée en faveur du sexe le plus profitable en terme de valeur sélective. Nos recherches concernent ces deux niveaux d’approches.
Biais de sex-ratio au niveau populationnel
Chez les insectes, notamment chez les parasitoïdes, les biais de sex-ratio au niveau populationnel sont assez fréquents. Dans son travail de thèse, M. Metzger étudie le biais de sex-ratio en faveur des femelles, observé en conditions naturelles chez Venturia canescens, en le comparant avec le modèle de la Local Mate Competition (Hamilton 1967, la condition où un nombre limité de femelles pondent leurs œufs dans un environnement clos) et avec le modèle alternatif de « female-constrained » (la condition où les femelles ont épuisé le stock de spermatozoïdes acquis lors des copulations et sont forcées à ne produire que des mâles, Godfray 1990).
Chez les vertébrés supérieurs par contre, les preuves d’un biais de la sex-ratio à l’échelle de la population restent rares. Nous avons montré que la sex-ratio secondaire (au sevrage) dans les portées de marmottes alpines est significativement biaisée en faveur des mâles (0,58). Ce biais peut s’expliquer par le modèle de la LRE (Local Resource Enhancement : le cas où des subordonnés restent dans le groupe familial, aident les parents à élever les jeunes et augmentent ainsi le succès reproducteur), dans la mesure où les subordonnés mâles peuvent être considérés comme des helpers (Allainé et Theuriau, 2004). L’aide fournie par les subordonnées mâles peut alors être considérée comme un remboursement de l’investissement parental, ce qui justifie leur production en excès.
Ajustement facultatif de la sex-ratio au niveau individuel
L’ajustement de la sex-ratio à l’échelle individuelle reste sujet à polémique pour deux raisons :
-
Les illustrations d’un tel ajustement ne montrent pas de patron consistant et sont controversées, en partie parce que les prédictions théoriques concernant la direction du biais attendu ne sont pas claires. Ainsi, le cerf en milieu forestier, soumis à une moindre intensité de la sélection sexuelle (Bonenfant et al. 2005), ne montre pas de variation d’allocation maternelle suivant le sexe des jeunes (Bonenfant et al. 2003). Même lorsque les conditions d’application du modèle de Trivers et Willard sont vérifiées, il faut en plus que les différences de qualité phénotypique entre femelles se traduisent en terme d’accès aux ressources. Ce n’est pas le cas chez le mouflon des Rocheuses et aucune variation de sex-ratio n’a pu être détectée en fonction de la qualité phénotypique de la mère (Blanchard et al., 2005). Enfin, lorsque l’importance des conditions initiales et la variance de valeur sélective sont plus influentes pour les femelles que pour les mâles, on peut assister à un renversement de situation et observer que les mères de meilleure qualité phénotypique allouent proportionnellement plus à leur fille qu’à leur fils (Hewison et al., 2005). Une prédiction claire quant à la direction de l’ajustement de la sex-ratio peut néanmoins être faite pour les espèces de vertébrés se reproduisant en coopération : la sex-ratio dans les portées devrait être biaisée en faveur des « helpers », lorsque ceux-ci sont absents des groupes familiaux. Cette prédiction est confirmée chez la marmotte alpine (Allainé 2004) : la sex-ratio dans les portées est biaisée en faveur des mâles (0,66) en absence de « helpers », mais équilibrée (0,49) en présence de « helpers », conduisant globalement au biais mâle observé à l’échelle de la population (Fig. 4).
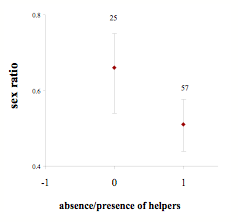
Figure 4 : Sex-ratio dans les portées de marmottes alpines quand les helpers sont absents (0) ou présents (1). La sex-ratio est la proportion de mâles. Les barres indiquent l’intervalle de confiance à 95%. Les nombres indiquent les nombres de portées considérées.
-
Les mécanismes de l’ajustement facultatif de la sex-ratio demeurent largement inconnus chez les vertébrés supérieurs. À partir de données récoltées sur le tahr en Nouvelle Zélande, nous avons montré que le mécanisme le plus probable pour une variation de sex-ratio en fonction de la qualité phénotypique des mères restait la mortalité différentielle favorisant les femelles in utero (Forsyth et al., 2004).
Des traits d’histoires de vie à la structuration des populations : aspects démographiques, spatiaux et génétiques
Démographie et variabilité environnementale
Bien que l’influence de la densité-dépendance sur la dynamique d’une seule population ait été largement étudiée, les effets de cette densité dépendance sur les systèmes hôtes parasitoïdes ont reçu peu d’attention. Après avoir montré expérimentalement que le parasitisme pouvait considérablement modifier l’effet de la compétition sur les hôtes (Bernstein et al., 2002, Fig 5), nous avons développé un modèle structuré en âge dans lequel la compétition intraspécifique est prise en compte selon une fonction densité dépendante flexible. Nous montrons que la présence du parasitoïde dans le système change les effets de la compétition intraspécifique entre hôtes sur la dynamique du système. En particulier, l’intensité de la compétition (c’est à dire, la position sur le continuum « contest-scramble »), qui joue un rôle crucial en dynamique de populations isolées, a un effet plus faible sur les populations sujettes au parasitisme. Nous montrons aussi que la stabilité du système est plus grande lorsque les hôtes sains sont plus susceptibles à la compétition que les parasités (Spataro et Bernstein, 2004).
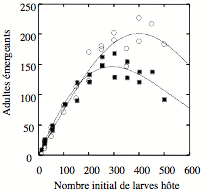
Figure 5 : Différences de susceptibilité à la compétition entre des larves saines du lépidoptère Ephestia kuehniella et des larves parasitées par le parasitoïde Venturia canescens. Le graphique montre le nombre d’adultes de parasitoïde (cercles) et d’hôte (carrés) en fonction du nombre initial de larves de chaque type (saines ou parasitées) en compétition. Les résultats montrent que les larves parasitées (et qui produiront des parasitoïdes adultes) sont moins susceptibles à la compétition (Bernstein et al., 2002). Spataro et Bernstein (2004) suggèrent que ce type de différences contribue à la stabilité du système formé par l’hôte et son ennemi naturel
Dans les populations de grands mammifères, les sources de variation environnementale telles que le climat et la densité de population interagissent pour déterminer le patron de variation des paramètres démographiques. Ces populations sont fortement structurées en âge et il est important de prendre en compte en particulier la différence entre adultes et vieux animaux pour obtenir une image fiable du fonctionnement de ces populations (Festa-Bianchet et al., 2003). La réponse des populations de grands mammifères aux facteurs de variation temporelle est aussi fortement structurée selon l’âge : la performance des juvéniles est ainsi toujours plus faible et plus sensible à la variation temporelle que celle des adultes (Gaillard et Yoccoz, 2003).
Parallèlement, le taux de croissance des populations de grands mammifères et par là, la valeur sélective des individus est plus sensible à un changement donné de survie adulte qu’au même changement de survie juvénile. On est donc en présence d’une relation négative entre variation relative des paramètres démographiques et leur impact potentiel sur la valeur sélective (Gaillard et Yoccoz, 2003). Dans ces conditions, le paramètre-clé de la dynamique des populations de grands mammifères ne correspond pas toujours au paramètre le plus influent potentiellement. Une étude comparative de quelques études de cas montre que ce paramètre-clé est contexte-dépendant et semble déterminé par le type de facteurs limitants (prédation versus ressources) et/ou le statut de la population (colonisation versus saturation) concernée (Coulson et al., 2005). Les variations inter-annuelles des conditions environnementales ne suffisent cependant pas à expliquer l’étendue de la variation des variations inter-individuelles des performances démographiques. Ainsi, même dans une population de cerfs norvégiens dans laquelle la variation inter-annuelle des poids de juvéniles est forte, une large composante de la variation inter-individuelle est due à des différences de performances maternelles (Loison et al., 2004). D’un autre coté, les femelles semblent avoir peu de latitude dans leur niveau d’investissement maternel (Loison et al., 2005) : dans des conditions environnementales défavorables, elles n’allouent pas plus d’énergie dans leur jeune, ce qui conduit à une forte variabilité de la survie juvénile.
Structures spatiale et génétique
Les individus sont distribués dans l’espace en fonction de la structure de l’habitat et du paysage, formant des sous-unités de populations plus ou moins discontinues. Nous avons cherché à mettre en évidence les différentes échelles de structuration spatiale de populations d’ongulés suivis sur le long terme (chamois, mouflons, chevreuils dans la réserve nationale de chasse et de faune sauvage des Bauges, chevreuils dans les sites de Chizé et de Trois Fontaines), et à identifier les causes de ces structures spatiales. L’étude intensive à long terme de la population de chevreuil de Chizé a permis de démontrer que (i) l’hétérogénéité spatiale de la qualité de l’habitat influence la masse corporelle des faons en hiver (Pettorelli et al., 2001), ainsi que les composantes estivale (Pettorelli et al., 2005a) et hivernale (Pettorelli et al., 2003a) de la survie de ces faons, (ii) ces influences sont détectables aussi bien à une échelle large (type de peuplement, quelques centaines d’hectares) que fine (domaine vital, quelques dizaines d’hectares) et sont toujours beaucoup plus marquées à une échelle fine, et (iii) ces influences spatiales interagissent avec les sources de variation environnementale et sont marquées seulement dans les mauvaises années. La présence d’espèces végétales préférées par les chevreuils (Pettorelli et al., 2003b, 2005b) et la qualité (teneur en azote) des plantes sont actuellement les meilleurs descripteurs de la qualité de l’habitat pour le chevreuil. Cependant, les données satellitaires permettant, entre autres, d’estimer la productivité primaire d’une zone donnée apparaissent prometteuses pour les analyses ultérieures (Pettorelli et al., 2005c). Ces recherches seront intégrées dans un contexte comparatif plus large. En effet, Hervé Fritz, qui a déposé une ATIPE dans notre laboratoire, travaille depuis de nombreuses années sur une thématique similaire à partir du modèle ongulés africains (Parc National de Hwange, Zimbabwe). Dans ce type d'écosystème, la pluviométrie annuelle plutôt que la sévérité hivernale ou la pluviométrie en été est le principal facteur limitant, et la réponse de la communauté d'ongulés pourrait varier avec ce changement de facteur limitant (Gaidet, 2005 thèse de doctorat). Une analyse comparative entre la variabilité spatio-temporelle du fonctionnement des communautés d'ongulés dans un écosystème de montagne (Les Bauges, programme de recherche d'Anne Loison) avec celle de cet écosystème tropical étudié par Hervé Fritz nous offre une opportunité unique d'appréhender les mécanismes de coexistence des espèces de grands herbivores dans des situations contrastées, et d'identifier par là les mécanismes généraux qui régissent cette coexistence.
6) Perspectives
Évolution des systèmes d’appariement et biais de sex-ratio
L’haplo-diploïdie caractéristique des hyménoptères parasitoïdes est source de conflits entre sexes. Ces conflits de choix de stratégie reproductive peuvent concerner: i) le nombre de partenaires sexuels ; ii) le nombre de spermatozoïdes transférés du mâle vers la femelle, iii) le sexe des descendants (i.e. la sex-ratio). L’intensité du conflit « d’intérêt » entre sexes concernant la sex-ratio dépend d’une part, du système d’accouplement (mating system) et d’autre part, des informations (internes et externes) que les femelles utilisent pour modifier la valeur de la sex-ratio. Notre objectif est de comprendre, en prenant l’exemple de Venturia canescens, les mécanismes et les informations impliqués dans ces conflits et d’en mesurer les conséquences évolutives en terme de nombre, sexe et qualité phénotypique des descendants (thèse de Marie Metzger).
L’existence de paternités hors couples (EPP) est maintenant largement documentée chez les vertébrés supérieurs. La polémique porte actuellement sur les causes évolutives de ces EPP, et deux grandes hypothèses s’affrontent : l’hypothèse des « bons gènes » et l’hypothèse de la « compatibilité génétique ». Nos résultats sur la marmotte alpine confortent la deuxième hypothèse. Notre premier objectif sera donc d’examiner la deuxième hypothèse plus en détail. De plus, peu de travaux concernent les conséquences des EPP en terme de fitness. Notre second objectif sera d’analyser les bénéfices, en terme de fitness pour les femelles, de la présence de jeunes issus de paternités hors couple.
Allocation optimale des ressources et variabilité environnementale
En continuité avec nos recherches passées, nous souhaitons développer un nouvel axe de recherches centré sur l’allocation optimale des ressources en fonction de la variabilité environnementale, du régime démographique et de l’intensité de la sélection sexuelle afin de mettre à l’épreuve les hypothèses théoriques de la biologie évolutive. Cet axe de recherche bénéficie maintenant d’un soutien fort grâce aux collaborations au niveau international avec la Norvège (GDRE soumis), le Canada et la Grande Bretagne, et présente ainsi une très forte faisabilité. À partir d’études de biologie de population complètes et à long terme, nous chercherons à identifier les covariations entre les traits d’histoire de vie classiquement étudiés mais aussi entre ces traits et les tactiques d’allocation des ressources, la variabilité environnementale, l’hétérogénéité spatiale, et l’intensité de la sélection sexuelle. Ceci devrait permettre de dépasser la vision démographique des stratégies évolutives pour aboutir à une vision biodémographique plus à même de permettre la mise à l’épreuve des théories de la biologie évolutive.
Les traits d’histoire de vie et la dynamique des systèmes hôte parasitoïde
Dans la période qui s’achève et dans des périodes précédentes nous avons fait des efforts pour introduire des descriptions réalistes des traits d’histoire de vie et du comportement dans les modèles de dynamique des systèmes hôte-parasitoïde. Nous comptons continuer dans cette voie, d’une part en apportant des connaissances sur les traits d’histoire de vie des parasitoïdes et de leurs hôtes et d’une autre part, en contribuant à la modélisation de la dynamique et de l’évolution de ces processus grâce à des développements comme la dynamique adaptative.
Analyse comparative des traits d’histoire de vie des punaises vecteurs de la maladie de Chagas
Malgré une littérature très abondante sur les punaises vecteurs de la maladie de Chagas, nous manquons d’analyses comparatives sur les traits d’histoire de vie des différentes espèces. Ainsi dans le cadre d’une collaboration franco-mexicano-argentine et un soutien du programme Ecos-Sud, nous proposons d’étudier l’évolution des traits d’histoire de vie et des caractères morphologiques chez ces punaises en tenant compte des relations phylogénétiques et des compromis évolutifs (trade-offs) notamment entre la dispersion et les autres traits d’histoire de vie. Notre approche associe une analyse des données de la littérature et celle de nos propres travaux expérimentaux. Ce programme implique une collaboration entre l’Université de Buenos-Aire, Argentine (J. Rabinovich), l’Université Autonome du Yucatan, Mexique (E. Dumonteil), notre département (F. Menu, C. Bernstein, E. Desouhant, D. Chessel et D. Pontier), l’Université de Perpignan (S. Gourbière) et l’Université de Tours (C. Lazzarri).
Relation entre les stratégies de ponte et les stratégies de diapause
Les stratégies de pontes chez les insectes ont été abordées essentiellement à partir d’une approche déterministe et indépendamment des stratégies de diapause. Cette approche n’a pas permis d’expliquer de nombreux exemples de variabilité dans le nombre d’œufs pondus par hôte. Chez le balanin de la châtaigne, les femelles ont adopté une stratégie de ponte mixte : elles pondent en moyenne 50% de leurs oeufs solitairement et 50% en multi-infestation (plus d’un oeuf par fruit). Pourtant, en raison d’un coût associé aux pontes solitaires inférieur à celui des multi-infestations, la théorie déterministe prédit une sélection des multi-infestations. Selon l’approche déterministe, la stratégie observée serait donc mal-adaptée. Ce paradoxe serait levé si la stratégie de ponte mixte observée permettait aux femelles de générer une variabilité dans la durée de diapause de leur descendance (via la relation entre la proportion de diapause prolongée et la quantité de ressources énergétiques des larves mise en évidence précédemment). Nous nous proposons de tester cette hypothèse (collaboration F. Menu et E. Desouhant).
Relations Interspécifiques
Chaque espèce interagit avec un ensemble de parasites, de prédateurs et de compétiteurs. Des coûts et des bénéfices sont associés à ces interactions, engendrant des pressions de sélection fortes qui peuvent influencer les structures génétiques des populations et les traits d’histoire de vie des espèces. Les effets de ces interactions peuvent varier en fonction des caractéristiques du milieu et dans le temps. La compréhension de ces interactions et de leur rôle dans l’évolution passée et future des espèces impliquées passe en particulier par une étude de leur cadre écologique.
1) Systèmes hôtes-parasites
L’étude des systèmes hôtes-parasites constitue un champ d’investigation d’importance croissante depuis deux décennies. Dans ce cadre, notre approche combine étroitement les travaux de terrain et la modélisation mathématique, ces composantes permettant une meilleure compréhension des dynamiques complexes des populations d’hôte(s) et de parasite(s).
Les modèles épidémiologiques actuels visent i) à comprendre les mécanismes à la base de la propagation observée d’une infection donnée et ii) à prédire son évolution dans les temps à venir, à échéance plus ou moins longue. Ce dernier point, d’un intérêt primordial pour la santé publique ou l’économie, reste encore difficile à préciser actuellement. Les principales sources d’hétérogénéité affectant la propagation des pathogènes sont i) l’hétérogénéité spatiale au sein des populations d’hôtes, ii) l’hétérogénéité comportementale des hôtes, iii) les différences de dynamique des populations (d’hôtes et/ou de pathogènes), iv) les stratégies de transmission et les différentes routes associées, ou encore v) le degré de protection immunitaire. Nous avons exploré l’impact de ces facteurs sur la dynamique et l’évolution des systèmes hôtes-parasites.
Hétérogénéité spatiale et dynamique des systèmes hôtes-parasites
De façon générale, l’hétérogénéité spatiale a un effet stabilisateur de la relation hôtes-parasites. Cependant, la modélisation et l’observation de populations spatialement hétérogènes, lorsqu’elles sont effectuées à une échelle vaste, ne permettent pas d’expliciter les mécanismes qui sont à l’origine de cet effet. Nous avons cherché à élucider les mécanismes de propagation et de persistance à différentes échelles.
Les modèles épidémiologiques actuels visent i) à comprendre les mécanismes à la base de la propagation observée d’une infection donnée et ii) à prédire son évolution dans les temps à venir, à échéance plus ou moins longue. Ce dernier point, d’un intérêt primordial pour la santé publique ou l’économie, reste encore difficile à préciser actuellement. Les principales sources d’hétérogénéité affectant la propagation des pathogènes sont i) l’hétérogénéité spatiale au sein des populations d’hôtes, ii) l’hétérogénéité comportementale des hôtes, iii) les différences de dynamique des populations (d’hôtes et/ou de pathogènes), iv) les stratégies de transmission et les différentes routes associées, ou encore v) le degré de protection immunitaire. Nous avons exploré l’impact de ces facteurs sur la dynamique et l’évolution des systèmes hôtes-parasites.
Hétérogénéité spatiale et dynamique des systèmes hôtes-parasites
De façon générale, l’hétérogénéité spatiale a un effet stabilisateur de la relation hôtes-parasites. Cependant, la modélisation et l’observation de populations spatialement hétérogènes, lorsqu’elles sont effectuées à une échelle vaste, ne permettent pas d’expliciter les mécanismes qui sont à l’origine de cet effet. Nous avons cherché à élucider les mécanismes de propagation et de persistance à différentes échelles.
À l’échelle d’un petit nombre de groupes, nous avons modélisé la propagation du Virus Leucémogène Félin (FeLV) dans des populations de chats en considérant la dispersion d’individus entre les populations. Pour cette infection, présente de manière hétérogène selon les populations de chats, l’objectif était de prédire l’effet de la dispersion des individus entre les populations (Fig. 6)). Le modèle prédit que la dispersion augmente la persistance virale, la fréquence d’infection et l’effet dépresseur de l’infection sur la population (Fromont et al. 2003). Ce modèle constitue un des premiers travaux où dispersion et densité-dépendance sont combinés pour prédire la propagation d’une infection.
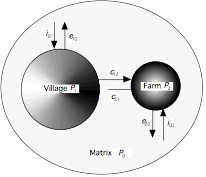
Figure 6 : Représentation des contacts entre populations dans le modèle de propagation du Virus leucémogène félin (FeLV) : les individus sont groupés dans des populations (villages ou fermes) et peuvent disperser d’une population à l’autre, mais aussi de ou vers la matrice, qui représente les chats errants dans les environs. Cette dispersion augmente la persistance du virus, la fréquence de l’infection et l’effet dépresseur du virus sur la taille des populations d’hôtes.
Lorsque de nombreux groupes sont en relation, les effets à l’échelle de l’ensemble d’une région peuvent émerger de la connexion entre populations locales. Nous étudions les mécanismes de transmission et de persistance de deux infections largement présentes dans les populations françaises de Sanglier (Sus scrofa) : la peste porcine classique et la brucellose. La propagation de la peste porcine est conditionnée localement par les effectifs et la dynamique de la population (Rossi et al., 2005a). Cependant, sa persistance à long terme est liée à la connectivité entre les populations locales d’une région (Rossi et al., 2005b). La brucellose au contraire semble pouvoir se maintenir localement, indépendamment de la dynamique de la population locale, mais sa distribution est influencée par la structure sociale et le mode d’appariement du Sanglier (thèse Rossi 2005).
Variabilité comportementale des hôtes, dynamique et évolution du mode de transmission des microparasites
La variabilité comportementale des hôtes se traduit par une hétérogénéité du risque d’infection et de contamination (Natoli et al., 2005) qui est déterminante dans la propagation et la persistance de la maladie. Le Virus d’Immunodéficience Féline (VIF) qui infecte les chats domestiques a été pris comme exemple.
Nous avons développé une approche qui couple un modèle d’évolution comportementale (basé sur la théorie des jeux) et un modèle épidémiologique déterministe en compartiments. Grâce à ce modèle, nous avons montré que les changements au niveau de l’accès aux ressources influent sur la fréquence des combats au sein des populations de chats et de ce fait sur la propagation du VIF. Ceci permet d’expliquer les différences de prévalence observées entre les populations rurales et urbaines (Bahi-Jaber et al., 2003). Selon le modèle, pour persister, le VIF doit être fréquemment réintroduit dans les populations. Or, en milieu rural, ce sont essentiellement les mâles dominants qui sont en contact avec des chats d’autres populations au moment de la reproduction. Un modèle couplant plusieurs populations de chats via ces mâles montre que la persistance du VIF est maximale pour la structure des contacts observée en milieu rural. Au contraire, en milieu urbain où les contacts entre populations sont très limités et concernent les femelles (Devillard et al., 2003b), la persistance du VIF est faible (thèse Bahi, 2003). Ces résultats montrent que les individus avec un comportement à risque plus marqué sont essentiels dans le processus de persistance du microparasite. Ceci explique probablement le mode de transmission du VIF que l’on observe dans les populations naturelles de chats. Mais cette stratégie n’est plus optimale dans le milieu urbain, très récent à l’échelle de l’évolution du système chat-VIF. La modification de la structure sociale et du patron de dispersion des chats en milieu urbain pourrait conduire à un mode de transmission permettant la propagation du VIF entre les populations de chats urbains via les femelles (thèse Bahi, 2003, Bahi et al., soumis).
Fonctionnement des populations et risque d’émergence
La néphropathie épidémique (NE) est une zoonose causée par le hantavirus Puumala, dont le réservoir est le campagnol roussâtre. Cette maladie affecte l’Europe du nord et du nord-ouest. En France la NE se cantonne au quart Nord-Est du pays, alors que le campagnol roussâtre est présent jusqu’à la méditerranée ce qui donne une situation propice à la recherche des mécanismes d’émergence de la maladie.
Les chutes brutales de densité du rongeur nous ont amené à poser la question de la persistance du virus lors de ces creux. Un premier travail de modélisation a permis de suggérer que le sol contaminé servait de réservoir au virus (Sauvage et al., 2003). Nos travaux ont aussi montré une augmentation simultanée de l’incidence de l’infection chez le rongeur et chez l’homme (Sauvage et al., 2002) qui n’était pas attendue, compte tenu du mode de transmission du virus à l’homme. Pour explorer ce phénomène, nous avons établi un modèle couplant la dynamique de la NE chez l’homme et celle de d’infection dans le réservoir campagnol (Fig. 7).
Le mécanisme proposé permet de comprendre les raisons pour lesquelles la répartition géographique des cas de NE ne recouvre pas entièrement celle des campagnols. Il implique également qu’un changement de la dynamique du rongeur (suite par exemple à des modifications environnementales) pourrait conduire à l’apparition de cas de NE dans des zones géographiques jusque là indemnes. Ce mécanisme d’apparition de la NE peut être généralisé à plusieurs autres maladies (Sauvage, thèse 2004, Sauvage et al., Soumis). Ce travail est réalisé en collaboration avec l’AFSSA (D. Augot, Nancy) et est soutenu par l’INSERM.
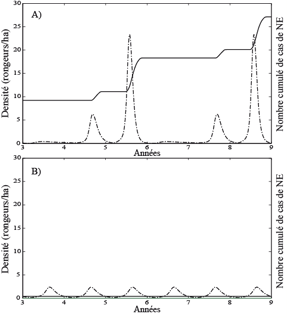
Figure 7 : Simulation de la dynamique de la néphropathie épidémique. Densité de campagnols en phase aiguë d’infection (courbe discontinue) et nombre total de cas humains de NE depuis l’introduction du virus (courbe continue), pour les deux types de dynamique démographique des populations de rongeurs. A) lorsque la population de campagnols exhibe des pics de densité élevée et B) lorsque la population réservoir est plus stable avec des maxima de densités intermédiaires.
De nombreux parasites possèdent un cycle complexe, notamment caractérisé par l’alternance de phases de reproduction sexuée et asexuée. L’importance relative de ces phases joue un rôle dans l’épidémiologie de ces parasites et dans leur évolution (Fig. 8).
Dans le cas du protozoaire Toxoplasma gondii, nous cherchons à estimer l’importance de la phase sexuée présente chez les félidés par rapport à la multiplication asexuée chez les autres hôtes, pour contribuer à l’estimation du risque d’infection chez l’homme. Dans le cadre d’un programme de recherche financé par l’Agence Française de Sécurité Sanitaire Environnementale (AFSSE), nous étudions l’hypothèse selon laquelle les modes de transmission majoritaires de la toxoplasmose diffèrent en fonction de l’environnement. Dans les populations de chats, nous avons montré que l’infection est variable entre populations mais aussi localement dans les populations et au cours du temps (Afonso et al., 2005). La densité, la structure des populations, le régime alimentaire et les caractéristiques physiques susceptibles de modifier la survie des formes environnementales de parasites peuvent influencer la dynamique de la toxoplasmose.
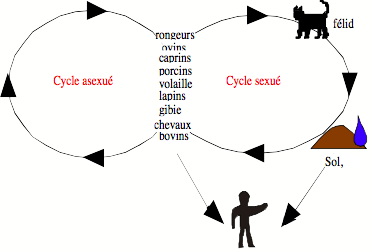
Figure 8 : Cycle du protozoaire Toxoplasma gondii. La multiplication sexuée a lieu chez un félidé, et les ookystes produits se développent dans le sol avant d’être ingérés par les hôtes intermédiaires. Les félins s’infectent en général par carnivorisme (cycle sexué). Cependant, le parasite peut persister pendant plusieurs générations d’hôtes intermédiaires par transmission verticale ou par carnivorisme (cycle asexué). L’importance relative de ces deux cycles varie en fonction des espèces présentes localement et de la survie des ookystes dans l’environnement. Pour l’homme, la part de risque due à la contamination par le sol et par les hôtes intermédiaires est attendue pour varier également.
Adaptation des Réseaux de Petri à la modélisation en épidémiologie
Les modèles en compartiments déterministes basés sur des équations différentielles sont bien adaptés à la prise en compte de la démographie, de la structure et de l’hétérogénéité de la population hôte. En revanche, ils sont inadaptés pour étudier l’effet de la variabilité stochastique sur la propagation de la maladie. De plus, ces modèles ne permettent pas de reproduire toute la complexité du processus de transmission.
Nous nous sommes donc orientés vers les Réseaux de Petri (RdP), un outil mathématique et graphique largement utilisé en ingénierie. Nous avons adapté ce formalisme à l’épidémiologie et montré que cette approche permettait de décrire de façon réaliste le processus de transmission de la maladie en tenant compte des aspects stochastiques impliqués et les caractéristiques individuelles (génétiques, histoire de vie par exemple) (Bahi, thèse 2003 ; Bahi-Jaber et Pontier, 2003). Ces caractéristiques font de cette approche un outil accessible aux biologistes leur permettant de façon rapide d’élaborer et d’analyser un modèle épidémiologique stochastique.
2) Coexistence d’espèces compétitrices
La célèbre question « Why are there so many species ? » reste une question centrale de l’écologie actuelle. Outre son intérêt en recherche fondamentale, les tentatives pour répondre à cette question se sont multipliées au cours des dix dernières années face à la demande sociétale de maintien de la biodiversité.
La diversité en espèces dans des écosystèmes locaux a toujours été discutée en relation avec la compétition inter-spécifique et de nombreux travaux théoriques et empiriques tentent de comprendre les mécanismes permettant à des espèces compétitrices de coexister localement. Le maintien local d’une population a souvent été envisagé comme étant dépendante du degré de compétition entre espèces pour des ressources limitantes. Les premières investigations théoriques traitant de ce sujet suggèrent que la compétition inter-specifique devrait conduire à la disparition locale des espèces les moins performantes (principe d’exclusion liée à la compétition). Paradoxalement, la coexistence d’espèces exploitant les mêmes ressources est très souvent décrite dans de nombreuses communautés et l’un des challenges actuels est de comprendre les mécanismes évolutifs permettant ce type de coexistence.
L’approche actuelle de la coexistence d’espèces compétitrices considère la multiplicité des facteurs pouvant agir sur la composition des communautés comme la prédation, le parasitisme, l’hétérogénéité spatiale et les fluctuations temporelles des ressources via des trade-offs inter-spécifiques. Dans ce cadre, différents travaux qui associent des approches complémentaires (modélisation, études de terrain et expérimentations en laboratoire) ont été réalisés au sein de notre UMR. Ces travaux portent en particulier sur la coexistence des modes de reproduction sexuée et asexuée, sur la coexistence d’espèces généraliste et spécialiste ou encore sur la coexistence d’espèces exploitant des ressources dont la disponibilité fluctue dans le temps de manière stochastique.
La coexistence des modes de reproduction sexuée et parthénogénétique
Comme nous l’avons vu par ailleurs mis à part les processus à long terme et en raison du coût supplémentaire que signifie la production de mâles, des lignées asexuées devraient rapidement exclure les compétiteurs sexués. Le problème de la coexistence des deux modes de reproduction, est, en beaucoup de points similaire à la coexistence de deux populations d’espèces en compétition pour des niches écologiques très proches. Pour étudier ce problème, nous avons construit un modèle largement inspiré du système formé par des populations sexuées et parthénogénétiques de V. canescens (voir plus haut). Ce modèle, qui considère des méta-populations sexuées et asexuées dans un environnement spatialement explicite, étudie l’interrelation entre l’hétérogéniété spatiale, la compétition entre populations et la dispersion des individus (Fig. 9). Les résultats (Amat, 2005) montrent que, dans un environnement fragmenté, la coexistence est favorisée si les individus sexués ont des distances de dispersion plus larges que celles de leurs compétiteurs asexuées. Ce modèle a inspiré des expériences de dispersion des deux modes de reproduction réalisées sur le terrain (Amat, 2005 ; Amat et al., soumis) et au laboratoire (experiences en moulin de vol, collaborations avec le Research Institute of Crop Production, République Tchèque et l’Institut Fédéral de Technologie de Zurich, Suisse).
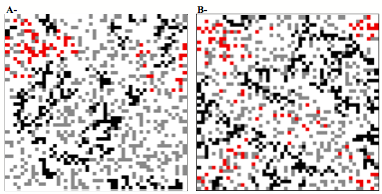
Figure 9 : Distribution spatiale des populations sexuées et asexuées après itération d’un modèle spatialement explicite pendant 40000 générations. L’environnement est fragmenté par la présence de sites indisponibles pour la reproduction (65%, figurés en blanc) aléatoirement répartis. Sont figurés en gris les sites disponibles, non colonisés. La coexistence des populations sexuées (en rouge) et asexuées (en noir) est favorisée si les individus sexués ont une distance de dispersion plus importante que celle de leurs compétiteurs asexués. (A) la distance de dispersion des individus asexués (distA) vaut 1 et celle des individus sexués (distS) vaut 2. (B) distA=1 and distS=5.
Stratégies d’exploitation des hôtes par les espèces spécialistes et généralistes
Un modèle a été développé pour formaliser et tester l’hypothèse selon laquelle l’évolution vers la spécialisation résulte de l’évitement de la compétition interspécifique. Ce modèle est inspiré du fonctionnement biologique d’oiseaux parasites de nichées originaires d’Amérique du sud. Ces parasites sont majoritairement des généralistes, chaque femelle pouvant pondre dans des nids d’un grand nombre d’espèces différentes (jusqu’à plus de 200 hôtes). Etre un parasite généraliste présente l’avantage de ne pas lier son succès reproducteur à celui de la population d’un hôte particulier. Pourtant, on connaît au moins un spécialiste, dans un système à trois espèces, l’hôte, le parasite spécialiste de cet hôte et un parasite généraliste. On peut donc se demander quelles sont les conditions qui permettent la persistance du spécialiste face au généraliste. Afin d’étudier l’influence de la compétition sur les populations couplées de l’hôte et du parasite spécialiste, nous avons construit un modèle de dynamique de population (Ney-Nifle et al., 2005). Les résultats montrent que le temps de production de chaque œuf, la destruction d’œufs par le parasite et la limitation du nombre total d’œufs qu’un hôte va tolérer dans son nid, jouent un rôle fondamental dans la stabilité du système et l’invasibilité par le spécialiste.
3) Perspectives
Les systèmes hôtes-parasites
L’étude des relations hôtes-parasites repose essentiellement sur une approche isolant chaque système microparasite-espèce(s) réservoir(s)/vectrice(s)-Homme. Nous défendons au contraire une approche comparative de plusieurs systèmes afin de mieux comprendre la diversité des facteurs et des mécanismes qui agissent seuls ou en synergie. Un aspect moins étudié que nous souhaitons développer est la relation qui peut exister entre les propriétés nosologiques d’un pathogène et la présence d’autres pathogènes et/ou la dynamique de la population hôte. Dans cette perspective, nous avons développé des réseaux de collaborations entre médecins, épidémiologistes, biologistes des populations, généticiens, virologues et mathématiciens autour de plusieurs pathologies qui peuvent affecter l’Homme et ont pour réservoir l’animal. L’approche interdisciplinaire associant la modélisation mathématique, le recueil des données sur le terrain, notamment à travers des suivis à long terme de populations animales, et le travail de laboratoire (identification du pathogène, typage des souches, typage génétique des hôtes) constitue le fil directeur transversal à l’ensemble des systèmes étudiés afin d’identifier des mécanismes fondamentaux de la relation d’un parasite avec son hôte, et ainsi de comprendre son évolution.
Coexistence d’espèces compétitrices
De nombreuses investigations théoriques récentes traitent des mécanismes évolutifs permettant la coexistence d’espèces compétitrices dans une même communauté. Les modèles biologiques avec lesquels nous travaillons et les outils méthodologiques que nous possédons nous permettent la mise à l’épreuve rigoureuse de ces développements conceptuels.
L’une des lacunes dans nos connaissances sur les processus de competition en conditions naturelles reside dans le fait que nous manquons d’expérimentations de terrain qui permettraient (1) de tester les prédictions des modèles, par exemple sur l’impact des fluctuations temporelles stochastiques et (2) d’étudier les compromis (“trade-offs”) et les déplacements de caractères liés à la compétition qui pourrait favoriser cette coexistence. Des travaux sur ces aspects seront développés avec deux modèles biologiques, des charançons du genre Curculio et les souches sexuées et parthénogénétiques de Venturia canescens.
Nous aborderons cette question chez le genre Curculio dont les espèces se développent aux dépens de fruits durs. Nous testerons l’hypothèse selon laquelle des variations temporelles stochastiques dans la production de fruits permet la coexistence d’espèces compétitrices via des trade-offs interspécifiques (e.g. dispersion vs fécondité). En parallèle, nous étudierons l’influence de la compétition interspécifique sur l’évolution de caractères phénotypiques (morphologie, traits d’histoire de vie).
Chez V. canescens nous concentrerons nos efforts dans l’étude des compromis liés à l’allocation de ressources pour le développement, la reproduction et la dispersion étant donné que les ressources mises à disposition par les hôtes sont limitées. Nous nous attendons à ce que la résolution de ces compromis diffère entre les deux modes de reproduction comme conséquences des différences entre leurs environnements préférentiels. Ces différences peuvent à leur tour se traduire dans des séparations plus profondes des niches. Nous étudierons en conditions de laboratoire et du terrain les différences en termes de i- longévité et fécondité, ii- de capacité dispersive dans l’espace et le temps iii- de comportement de recherche d’hôtes et de nourriture.
Dans les deux cas le travail combinera des travaux de terrain, d’écophysiologie et de modélisation mathématique.
Comportement individuel et processus populationnels
Le comportement permet des réponses rapides et flexibles de la part des animaux, à des changements de leur milieu naturel. Pour cette raison, le comportement joue un rôle particulièrement important dans l’adaptation des organismes à l’environnement. D’autres aspects ont participé aux succès des études comportementales. La relative facilité pour l’observation des processus et pour l’expérimentation au laboratoire et sur le terrain a permis l’élaboration d’un cadre conceptuel évolutif clair et la mise à l’épreuve rigoureuse des développements théoriques. Tous ces facteurs ont contribué à un développement assez équilibré des aspects théoriques et expérimentaux combinant les approches évolutives avec l’étude des mécanismes mis en jeu.
Une bonne partie de nos efforts est liée à l’étude de la dispersion et du choix de l’habitat. L’évolution de la coopération sujet, à la fois classique et centrale en écologie évolutive fait aussi partie de nos centre d’intérêts. Finalement, si l’influence de la variance dans les processus n’est pas oubliée en écologie du comportement, un travail sur les stratégies de capture par les araignées rappelle l’impact évolutif que peuvent avoir les événements rares.
1) Coopération
Un mammifère coopératif : la marmotte
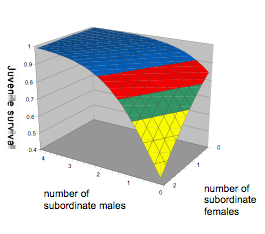
Figure 10 : Survie des marmottons pendant l’hiver en fonction du nombre de femelles et de mâles subordonnés dans le groupe. Légende des couleurs: bleue = s > 0.9 (nombre d’observations dans cette zone: n = 54); rouge = 0.8 < s <0.9 (n = 76); vert = 0.7 < s < 0.8 (n = 14); jaune = 0.4 < s < 0.7 (n = 17).
L’évolution de la reproduction en coopération (« cooperative breeding ») reste un sujet largement débattu. En particulier, les conséquences de la présence d’aides (« helpers ») sur la fitness des parents et des helpers eux mêmes, restent encore controversées.
Nos travaux sur la marmotte alpine ont contribué à montrer que cette espèce se reproduit en coopération (Allainé et Theuriau, 2004), les subordonnés mâles ayant le rôle de helpers. La survie des marmottons pendant l’hibernation augmente avec le nombre de subordonnés mâles présents dans l’hibernaculum, qui participeraient activement ou passivement à la production de chaleur. Il ne s’agit pas simplement d’un effet de la taille du groupe d’hibernation puisque la présence de subordonnés femelles diminue la survie juvénile pendant l’hiver (Fig. 10). Seuls les subordonnés mâles participeraient (activement ou passivement) à la production de chaleur alors que les femelles entreraient en compétition avec les marmottons pour la thermoregulation.
La présence de helpers représente aussi un coût pour le mâle dominant, et un modèle d’optimisation a permis de montrer que du point de vue de la fitness du mâle, il existe un nombre optimal de helpers dans les groupes familiaux (Allainé et Theuriau 2004).
2) Dispersion et colonisation
Les règles de dispersion chez un insecte parasitoïde
Bien que la dispersion soit un trait d’histoire de vie ayant d’importantes conséquences en termes évolutifs et écologiques (par exemple dans le contexte du contrôle biologique), notre connaissance de la dispersion chez les insectes parasitoïdes reste encore très limitée. Nous avons étudié les règles de navigation que le parasitoïde Venturia canescens emploie pendant sa migration entre populations d’hôtes en conditions naturelles (Desouhant et al., 2003). Les résultats montrent que les parasitoïdes dispersent rapidement (une heure pour une surface d’un hectare), évitent les espaces ouverts et ensoleillés et se dirigent vers la végétation et les endroits ombragés. Amat (thèse, 2005) a montré que les femelles sexuées et asexuées (cf section sur traits d’histoire de vie) emploient les mêmes règles de dispersion. Ces travaux se poursuivent par des expériences de laboratoire réalisées en collaboration avec le Research Institute of Crop Production, République Tchèque l’Institut Fédéral de Technologie de Zurich, Suisse et l’Université de Brême, Allemagne
Dispersion et colonisation chez la punaise Triatoma dimidiata, vecteur de la maladie de Chagas dans la péninsule du Yucatan au Mexique.
La maladie de Chagas transmise par Trypanosoma cruzi est responsable de millions de morts en Amérique latine. La seule stratégie disponible pour lutter contre cette maladie est le contrôle des vecteurs, des triatomines. L’optimisation de ce contrôle nécessite une connaissance de la dispersion spatiale de ces insectes et de la colonisation des zones habitées. Nous sommes engagés dans l’étude de la dynamique de dispersion de Triatoma dimidiata dans la péninsule du Yucatan au Mexique (collaboration avec de chercheurs mexicains et le Ministère de la Santé mexicain).
Nous avons montré que le taux d’infection des punaises par T. cruzi était élevé (34% en moyenne) (Dumonteil et al. 2002) et que contrairement à ce que l’on a observé dans d’autres pays d’Amérique latine, il n’existe pas de réelles populations domestiques. Les punaises ne sont présentes à forte densité dans les structures humaines que pendant la saison sèche (environ 3 mois) puis leur effectif chute drastiquement (Fig. 11).
Les résultats indiquent une dispersion saisonnière des zones sylvestres vers les habitations dont l’origine est encore inconnue. Un modèle mathématique (Gourbiere et al. soumis) suggère que les punaises qui arrivent dans les maisons durant la saison sèche n’arrivent pas à coloniser ces lieux en raison d’une reproduction insuffisante dans les habitations. Une analyse moléculaire est en cours (thèse en cotutelle de V. Payet) pour étudier si les insectes domestiques sont proches génétiquement des insectes sylvestres.
L’absence de populations domestiques indépendantes des populations sylvestres est à prendre en compte dans l’élaboration des stratégies de lutte contre les punaises. En effet un traitement insecticide local est inefficace à moyen terme en raison de la dispersion saisonnière des punaises des zones sylvestres vers les habitations (Dumonteil et al., 2004).
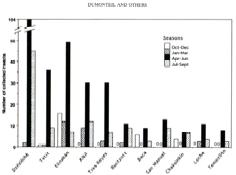
Figure 11 : Variation saisonnière de l’effectif total des populations de punaise Triatoma dimidiata, vecteur de la maladie de Chagas dans différents villages de la péninsule du Yucatan au Mexique (Dumonteil et al., 2002). Quatre périodes de l’année ont été consideré. Le nombre d’insectes collectés varie significativement selon la période. Très peu de punaises ont été trouvées dans les maisons entre octobre et mars. Des effectifs élevés sont caractéristiques de la période avril-juin. Les effectifs diminuent ensuite en juillet-septembre. Des données complémentaires suggèrent une invasion saisonnière des maisons via les zones sauvages environnantes (Dumonteil et al., 2004).
Patron de dispersion et structure des populations chez des mamifères
À partir d’une approche interspécifique, nous avons pu mettre en évidence une relation entre le niveau de complexité sociale et l’amplitude du biais sexuel de dispersion chez les rongeurs sciuridés polygynes (Devillard et al., 2004a). À partir d’une analyse comparative phylogénétique chez les rongeurs sciuridés polygynes, nous avons montré, en accord avec le modèle de Perrin et Goudet (2001), que l’amplitude de la différence entre les taux de dispersion mâle et femelle augmentait avec le niveau de socialité. Ces résultats vont dans le sens d’un rôle important de l’évitement de la consanguinité comme facteur évolutif de la dispersion natale chez les mammifères les plus sociaux.
Toutefois le patron de dispersion peut être extrêmement variable entre populations d’une même espèce, comme c’est le cas chez le chat domestique errant (Felis catus). Nous avons mis en évidence que la dispersion des chats en dehors de la population urbaine de Lyon était faible et principalement réalisée par des femelles entre 1 et 3 ans (Devillard et al., 2004b). Ce patron de dispersion est contrasté avec celui mis en évidence entre les groupes sociaux au sein de cette même population urbaine où les mâles se dispersaient plus que les femelles (Devillard et al., 2003). Dans la population introduite sur l’archipel des Kerguelen en milieu sub-antarctique, le patron de dispersion semble biaisé en faveur des femelles (Devillard et al. soumis, Devillard et Pontier, soumis). L’analyse des différentes populations suggère que le patron de dispersion dans les populations de chats errants co-varie avec la densité, le système d’appariement, et les structures sociale et spatiale, et ce, en accord avec les principales hypothèses théoriques au sujet des patrons de dispersion.
3) Information et apprentissage
Le comportement adapté des animaux dépend en grande partie de la capacité de perception des organismes, des différents types d’information que l’animal recueille, de l’intégration de ces informations et des mécanismes qui transforment l’information en actions (un comportement, une réponse physiologique, etc.). Selon la façon dont elle est obtenue, l’information sur les caractéristiques de l’environnement est souvent classifiée en «information publique » ou « information privée ».
L’information est dite publique quand elle est obtenue par l’observation du comportement et/ou le succès des congénères (en termes de succès reproducteur, obtention de ressources,…). L’information « privée » est obtenue individuellement, à partir de l’expérience de chaque animal. L’information « publique » se caractérise par l’efficacité de l’échantillonnage qu’elle permet mais aussi par un délai entre la récolte de l’information, et son utilisation. Pour cette raison, elle est plus souvent observée chez des animaux dont l’espérance de vie est de plus d’une année. L’information « privée » dépend d’une mise à jour permanente de l’évaluation, faite par chaque individu, des conditions de l’environnement et permet la simultanéité entre la collecte d’information et l’obtention de ressources. Elle est souvent observée chez les animaux de vie plus courte comme les insectes (van Alphen et al., 2003).
Information publique et sélection de l’habitat de reproduction
Un travail théorique réalisé avant l’incorporation de B. Doligez à notre laboratoire (Doligez et al. 2003) a confronté, en termes évolutifs (ESS) différentes stratégies d’obtention de l’information pour le choix de l’habitat de reproduction. Les résultats montrent que l’utilisation de l’information publique est favorisée quand l’environnement est suffisamment prévisible. En accord avec ces prédictions, des travaux expérimentaux et observationnels (Doligez et al., 2002, 2004), ont montré, chez une espèce qui se reproduit dans un environnement prédictible dans le temps, l’emploi de l’information publique dans le choix de l’habitat reproductif pour la saison de reproduction suivante.
Information privée et exploitation d’agrégats d’hôtes et de nourriture chez les insectes parasitoïdes
La vie adulte des parasitoïdes est en général assez courte. En conséquence une stratégie qui permet l’obtention de l’information simultanément à la collecte des ressources semble particulièrement appropriée. Néanmoins, l’utilisation d’information publique n’est pas absente chez les insectes. Ainsi V. canescens montre la capacité à discriminer à distance les sites de reproduction fortement exploités par des congénères (Castelo et al., 2003).
Lors de l’exploitation d’agrégats d’hôtes, le parasitoïde V. canescens emploie de l’information privée et intègre au moins trois sources d’information : le temps depuis l’arrivé à l’agrégat, la concentration des substances chimiques déposées par les hôtes et le nombre de pontes réalisés. Dans une revue (van Alphen et al. 2003), nous avons analysé la diversité de stratégies employées par les parasitoïdes. Cette revue suggère que la façon dont cette information est employée doit dépendre de la fiabilité de l’information disponible pour les parasitoïdes concernant la richesse en hôtes (Fig. 12). Nous avons formalisé cette idée grâce à un modèle basé sur l’individu et spatialement explicite (Spataro et Bernstein, soumis).
Figure 12 : Ces modèles graphiques illustrent deux stratégies d’abandon d’un patch selon la fiabilité de l’information obtenue par le parasitoïde en arrivant sur le patch. En arrivant l’animal a une tendance initiale à rester, cette tendance peut être (B) ou non (A) proportionnelle à la richesse du patch, cette tendance diminue avec le temps. (A) l’animal est incapable de discerner entre les patches riches et pauvres. C’est le nombre de pontes (indiquées par des flèches) qui révèle la richesse du patch : une diminution de la probabilité de quitter le patch comme conséquence de chaque ponte assure la meilleure exploitation du patch riche. (B), en arrivant l’animal obtient une indication fiable de la richesse du patch (H1 ou H2). Dans ce cas, les pontes sont une indication que le patch s’appauvrie progressivement, la tendance à partir doit augmenter à chaque ponte.
4) L’obtention de ressources
L’effet « jackpot » chez les araignées
Les événements rares et positifs sont souvent négligés en écologie du comportement du fait de leur faible fréquence et de la difficulté d’en estimer les conséquences en terme de fitness. L’effet que de tels événements rares peuvent avoir en termes de fitness a été clairement démontré par Venner et Casas (2005). Ces auteurs ont étudié la taille des toiles d’une araignée orbitèle (Zygiella x-notata) en relation avec la capture exceptionnelle de grosses proies. Ils ont exploré, à l’aide d’un modèle énergétique stochastique, les conséquences en termes de fitness de l’augmentation de la taille des toiles, en relation avec la capture des grosses proies. Les résultats montrent que ces proies ne sont qu’exceptionnellement capturées et les simulations suggèrent que les toiles d’araignées sont conçues pour capturer de rares et grosses proies (Fig. 13).
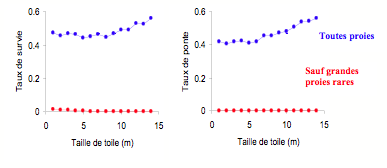
Figure 13 : Influence de la capture exceptionnelle de grandes proies chez l’araignée (Zygiella x-notata) : résultats d’un modèle de simulation. Taux de survie et de ponte en fonction de la taille des toiles en considérant (en bleu) toutes les proies attrappées par les araignées ou (en rouge) en excluant les proies de taille exceptionnelle. Les résultats montrent que les araignées ne peuvent ni survivre ni se reproduire sans capturer de grosses proies et que l’augmentation de la taille des pièges augmente à long terme la survie et la fécondité des araignées grâce à ces grosses proies rares.
Le gardiennage de ressources
Les stratégies de gardiennage précopulatoire du partenaire sexuel sont encore peu explorées à travers des expérimentations de terrain. Bel-Venner et Venner (2005) ont testé, chez l’araignée Zygiella x-notata, l’hypothèse que la stratégie de gardiennage devrait fortement dépendre du niveau de compétitivité des mâles. Le suivi longitudinal d’une population naturelle couplé à une expérience de terrain, a montré que, conformément aux prédictions théoriques, les grands mâles (plus compétitifs) délogeaient des gardiens déjà en place plutôt que de garder des femelles seules, qu’ils entraient plus tardivement en gardiennage, et qu’ils obtenaient un succès de gardiennage plus élevé.
5) Perspectives
Coopération
Les bénéfices de l’aide pour les dominants et pour les « helpers » sont bien illustrés, mais les coûts de l’aide pour les « helpers » restent sujet à débat. Notre perspective à court terme sera donc d’identifier le coût de l’aide pour les « helpers » chez la marmotte alpine. A plus long terme, il faudra déterminer si cette aide correspond à un comportement actif ou au sous-produit d’un comportement ayant une autre fonction (par exemple réveil pour la remise en activité des fonctions de reproduction).
Déterminisme de la dispersion chez les oiseaux
La dispersion a longtemps été considérée comme un trait fixé génétiquement, indépendant des conditions environnementales. Cependant, les travaux empiriques ont montré que les comportements de dispersion sont sous l’influence de nombreux facteurs internes aux individus (facteurs génétiques, condition individuelle, histoire personnelle) et externes (habitat, environnement social). Pour explorer la façon dont différents facteurs interagissent pour déterminer le comportement de dispersion, une expérience à long terme a été lancée en 2005 par Blandine Doligez sur le gobe-mouche à collier et la mésange charbonnière (expérience réalisée en Suède en collaboration avec des chercheurs de ce pays, PICS n° 3054 avec la Suède). Ce travail a comme objectif de distinguer chez ces espèces, l’influence des facteurs précoces (facteurs génétiques, effets maternels) de celle de facteurs tardifs (effets parentaux et environnements tardifs). L’un des aspect qui sera consideré est la question de comment différentes sources d’informations sont-elles intégrées par les individus, en particulier comment des informations contradictoires sont-elles gérées ?
Information et obtention de ressources chez les insectes parasitoïdes
Des résultats récents que nous avons obtenus montrent une capacité encore plus grande pour intégrer de l’information chez V. canescens que ce qui est décrit dans les pages antérieures. Ainsi, lors de la visite d’un agrégat d’hôtes, en plus des variables déjà mentionnées, V. canescens intègre l’information de ses propres ressources métaboliques et de la présence ou absence de nourriture dans le voisinage (Thèse de P. Lucchetta, en cours, collaboration avec E. Wajnberg). L’un des aspects que nous envisageons d’étudier ce sont les indices que V. canescens emploie pour repérer différentes ressources. Ainsi, nous sommes en train d’évaluer la capacité de ce parasitoïde à associer différentes couleurs à chaque ressource (travail en collaboration avec M. Théry, Muséum d’histoire Naturelle) et l’apprentissage contexte dépendant (collaboration avec A. Kacelnik Université d’Oxford). L’un des aspect fondamentaux de ce travail est d’identifier les ressources alimentaires réellement employées par cet insecte dans des conditions naturelles (collaboration avec D. Giron, Université de Tours, financement GDR 2155, Ecologie Comportementale).
L’exploitation de patches d’hôtes par des groupes de congénères chez V. canescens
L’étude du comportement de recherche d’hôtes par des groupes de congénères dans un système multi-patch (« Social foraging ») est un aspect fondamental de l’écologie du comportement des parasitoïdes que jusqu’à très récemment nous étions incapables d’aborder correctement. Néanmoins, le travail de Mabel Guerra (stage de master 2) a permis d’identifier les conditions expérimentales dans lesquelles les guêpes montraient un comportement stable vis-à-vis de ses congénères. Les résultats obtenus on permis d’identifier les conditions expérimentales adéquates et ouvrent la voie à une étude plus approfondie de ce sujet. Lors de cette étude nous combinerons les aspects théoriques et expérimentaux sur la distribution de congénères dans un environnement hétérogène dans des conditions où les parasitoïdes défendent activement les ressources déjà acquises (c’est-à-dire des hôtes qu’ils ont déjà parasité).
Gestion et conservation de la biodiversité
Les travaux récents en biologie des populations et en écologie évolutive, y compris dans ce laboratoire, ouvrent des perspectives pour améliorer la gestion et la conservation des espèces. Plusieurs travaux montrent que la seule prise en compte de la démographie et de la structure en âge de la population peut conduire à une mauvaise compréhension de son fonctionnement et à proposer des mesures inappropriées. La meilleure compréhension de l’écologie des populations permet au contraire de proposer des pistes de gestion qui intègrent la complexité de leur fonctionnement.
Notre laboratoire, sans être un acteur direct de la gestion, travaille en étroite collaboration avec plusieurs organismes chargés de mettre en place les mesures concrètes de gestion et de conservation : l’Office National de la Chasse et de la Faune Sauvage, l’Institut National de la Recherche Agronomique, l’Institut Paul-Emile Victor, Agence Française de Sécurité Sanitaire et Alimentaire ou les parcs nationaux. Ces collaborations visent à retranscrire dans le cadre de la gestion les connaissances acquises sur le fonctionnement des populations. Elles se traduisent concrètement par la mise en place par ces organismes de nouveaux outils de gestion. Ces outils doivent intégrer d’une part une meilleure prise en compte des traits d’histoire de vie des espèces, mais aussi les autres espèces en interaction avec les espèces visées, et le potentiel évolutif des populations en réponse aux mesures appliquées.
1) Gestion des populations
Concernant la gestion des populations, les recherches du laboratoire concernent l’élaboration d’outils de diagnostic plus pertinents que ceux classiquement utilisés, afin d’aider les gestionnaires dans leur prise de décision.Développement de méthodes de diagnostic des populations
Nos recherches sur la dynamique des populations d’ongulés sont conduites en étroite collaboration avec l’Office National de la Chasse (convention d’étude liant notre laboratoire et cet organisme, ininterrompue depuis 1989). Depuis plusieurs années, nos travaux ont montré que les méthodes classiques de comptages (dénombrement exhaustif ou par zone-échantillon) ne constituent pas une information pertinente pour l’établissement des règles de gestion (Gaillard et al., 2003a). Dans ces conditions, le suivi d’indicateurs de changements écologiques tels que le nombre d’animaux observés par kilomètre parcouru (Maillard et al., 2002), la masse corporelle des animaux (Bourgarel et al., 2002 sur l’impala ; Bonenfant et al., 2003 sur le cerf ; Kjellander et al. 2005 et Pettorelli et al., 2005 sur le chevreuil), le nombre de jeunes produits par femelle individualisée (Bonenfant et al., 2005 sur le cerf) ou l’impact des animaux sur la végétation constituent des mesures fiables des changements opérant dans la relation animal-habitat.
Développement de modèles de gestion
Les modèles de gestion des populations de grands mammifères reposent le plus souvent sur un modèle global de densité-dépendance (modèle logistique) et ne prennent pas en compte l’existence de structurations pourtant fortes dans les populations de ces espèces. Quatre structurations principales peuvent être reconnues à partir de nos travaux.
Les populations d’espèces de vertébrés de grande taille sont itéropares et fortement structurées en âge. Nous avons pu montrer que la distinction entre juvéniles et adultes n’était pas suffisante pour rendre compte du fonctionnement des populations d’ongulés. Il est en effet important de distinguer les adultes dans la force de l’âge des vieux individus sénescents. La sénescence est très répandue dans les populations de grands herbivores (Gaillard et al., 2003b), et conduit à détecter de la densité-dépendance dans la survie là où elle est absente (Festa-Bianchet et al., 2003).
Les différences de traits d’histoire de vie entre sexes souvent marquées dans les populations d’espèces dimorphiques et polygynes comme les grands herbivores (Toïgo et Gaillard, 2003 sur la survie adulte) peuvent également confondre l’étude du fonctionnement des populations. Des modèles structurés selon le sexe sont donc requis (Gaillard et al., 2003a).
Les conditions environnementales de l’année de naissance ayant des influences durables sur la performance des grands herbivores (Gaillard et al., 2003c), il est nécessaire de tenir compte de ces effets de cohortes, particulièrement pour les opérations de repeuplement ou de renforcement de populations (Gaillard et al., 2003a).
Enfin, il convient d’inclure l’hétérogénéité spatiale de la qualité de l’habitat dans la gestion pour éviter un prélèvement inadéquat par rapport à l’objectif de gestion (Pettorelli et al., 2002). Il faut aussi ajouter la composante financière qui est de plus en plus au centre des discussions lors de la définition des objectifs de gestion. Nous avons réalisé un premier modèle de ce type pour le chamois dans les Alpes (Skonhoft et al., 2002). Même si les estimations de nombreux paramètres restaient encore très approximatives, notre modèle indique qu’une optimisation bioéconomique est possible.
2) Gestion, conservation dans le cadre des relations interspécifiques
La gestion des populations peut nécessiter de prendre en compte les conséquences des mesures appliquées dans une population sur les espèces en interaction, notamment les proies ou les prédateurs. Dans les systèmes hôtes-parasites, la limitation de l’impact du parasite est même un objectif sous-jacent à la plupart des études (voir les cas de la myxomatose ou de la maladie de Chagas pour lesquels la vaccination ou le contrôle des vecteurs sont recherchés). Les mesures appliquées peuvent avoir aussi des effets non désirés. Des modèles sont alors nécessaires pour comprendre les facteurs clé en jeu et prédire la dynamique possible des systèmes sous l’effet des mesures de gestion proposées.
Effet de la dynamique de population et de la structure spatiale sur l’effet des mesures de gestion
Les principales mesures prophylactiques proposées pour limiter ou éradiquer une infection sont la vaccination et l’élimination des individus infectés. Ces mesures interfèrent avec la propagation de l’infection ainsi qu’avec la dynamique de la population. Il est donc nécessaire de prendre en compte l’action des mesures de gestion sur cette population. Dans le cas du Virus Leucémogène Félin (FeLV), nous avons étudié l’effet de la vaccination et de l’euthanasie des animaux infectés (Fig. 14).
Figure 14 : Prédiction de l’effet de la vaccination (à gauche) et de l’élimination des chats infectés par le virus leucémogène félin (à droite) appliquées dans une grande population de chats en contact avec une petite population. Chacune des mesures limite la prévalence globale (Poverall) et la prévalence dans la grande population (P1). Cependant, la prévalence dans la petite population (P2) augmente avec le taux de prophylaxie. Quel que soit le taux pratiqué, l’élimination ne permet jamais l’extinction du virus
Le modèle mis en place pour analyser la propagation de l’infection (Fromont et al. 2003) a été réanalysé en considérant spécifiquement le cas où parmi deux populations de chats, une seule est accessible à la prophylaxie. Ce modèle prédit que la vaccination est plus efficace que l’euthanasie. Cependant, les deux méthodes ont des effets inattendus : la prophylaxie appliquée à un taux réaliste dans une des populations peut avoir pour effet d’augmenter la propagation virale dans l’autre population. Ces résultats peuvent avoir des conséquences importantes lorsque des populations sauvages et domestiques échangent des agents pathogènes.
Hétérogénéités spatiales de la distribution des prédateurs en fonction de celles des proies et conservation des oiseaux en milieu insulaire
En général, les programmes d’éradication des espèces invasives en milieu insulaire n’ont concerné que les îles de petite taille (quelques km2). Les études de modélisation des interactions prédateur-proie se sont souvent intéressées à un système homogène dans le temps et dans l’espace. Or les patrons spatiaux qui émergent à partir des interactions localisées influencent fortement la dynamique des processus écologiques et épidémiologiques.
À l’image de la Grande Terre (3200 km2) de l’archipel des Kerguelen, les grandes îles sont souvent hétérogènes quant à la distribution des espèces, notamment indigènes (oiseaux marins) et introduites (chats, lapins, rongeurs).
La population de chats est morcelée en plusieurs sous-populations probablement interconnectées (Pontier et al., 2005), mais ayant des dynamiques locales différentes en raison de la disponibilité variable des proies dans le temps et dans l’espace (Pontier et al., 2002 ; Devillard (thèse, 2004) ; Devillard et al., soumis). À partir de modèles mathématiques, il apparaît que le système présente une diversité de comportements en fonction de la nature des hétérogénéités spatiales introduites (Gaucel et Pontier 2005 ; Gaucel et al., 2005). Ces résultats permettent de proposer des modalités de gestion et de conservation des espèces d’oiseaux marins menacés par la présence du chat, lorsque l’éradication de ce dernier n’est pas envisageable.
3) Gestion et évolution
Les mesures de gestion représentent aussi des forces sélectives qui conduisent les populations à évoluer. Nous étudions les effets de ces changements évolutifs chez les insectes où cette évolution est classiquement observée, mais nous montrons également de tels changements chez les Mammifères.
Évolution de la résistance aux insecticides et lutte biologique chez les insectes
Les insecticides ont des effets sur les écosystèmes beaucoup plus complexes que la simple diminution des effectifs des ravageurs. D’une part, l’action des insecticides sur des espèces non ciblées (souvent des auxiliaires de l’agriculture) génère des processus qui peuvent augmenter les effectifs et par conséquent, la dépendance aux insecticides pour la protection des cultures. Un second processus vient s’ajouter : l’apparition de résistances aux insecticides. Cette résistance permet de mieux tolérer la toxicité de l’environnement, mais à un coût énergétique qui se traduit par des effets délétères sur la reproduction ou le parasitisme. La détoxification a alors des conséquences à la fois dynamiques et évolutives qui sont intimement interdépendantes : la dynamique des populations fixe les conditions dans lesquelles la sélection naturelle opère, et est à son tour modifiée par le jeu des phénotypes évolutifs. Les travaux réalisés en collaboration avec A. Bernusset puis T. van Dooren (Université de Leiden, Pays Bas) ont montré que les processus évolutifs liés à l’effet des insecticides peuvent fondamentalement altérer la dynamique des populations et conduire à des effectifs cycliques dans le temps. Les résultats suggèrent aussi que lorsque la résistance affecte le comportement des hôtes et des parasitoïdes, elle a un effet plus déstabilisant que quand elle n’altère que des paramètres démographiques.
Modèle d’évolution des caractères morphologiques des ongulés en réponse à une chasse sélective
Dans la plupart des régions du globe, les populations de grands herbivores sont principalement limitées par la chasse. Cette chasse est le plus souvent sélective. Par exemple, beaucoup de populations d’ongulés très dimorphiques sont soumises à une chasse au trophée centrée sur les mâles les plus développés. Les effets pervers d’une telle sélection ont été démontrés sur le mouflon des Rocheuses en Alberta, Canada. Une étude en cours (thèse de M. Garel) a montré des effets similaires dans les populations de mouflons en France. Après de nombreuses années de chasse au trophée, la taille des cornes et la masse corporelle des agneaux ont ainsi diminué fortement, entraînant des performances démographiques plus faibles, et donc une stabilisation de l’abondance de la population.
4) Perspectives
Gestion des populations de mammifères
En ce qui concerne la gestion des populations de grands mammifères les enjeux de gestion ne sont plus limités au suivi d’unités de gestion particulières, mais nécessitent de passer à une échelle plus large tel que le département ou l’écosystème. Ce changement d’échelle de suivi appelle au développement d’outils de gestion nouveaux. Nous envisageons donc, en étroite collaboration avec des départements volontaires (Loire, Charente et Tarn en particulier) de tester la transposition d’indicateurs de changements écologiques validés au niveau de l’unité de gestion à l’échelle d’un département. Actuellement, le gestionnaire acquiert un ensemble de réponses d’indicateurs variés qui ne sont pas toujours faciles à interpréter. Un second objectif également de nature méthodologique consistera à quantifier la réponse des différents indicateurs dans une même mesure globale. Finalement nous visons à intégrer plus directement la composante qualité de l’habitat dans la gestion des populations d’ongulés. L’analyse des données sur l’occupation de l’espace par les ongulés dans différentes populations de plusieurs espèces permettra d’identifier les composantes de l’habitat les plus influentes dans chacun des cas. Toutes ces recherches seront effectuées en relation étroite avec l’Office National de la Chasse et de la Faune Sauvage.
La récente création de l’Unité Suivi Sanitaire de la Faune au sein de l’Office National de Chasse et de la Faune Sauvage témoigne de la prise de conscience de la nécessité d’intégrer à la gestion des populations, les effets des prédateurs, des pathogènes et des compétiteurs. Nos travaux montrent qu’il est possible de proposer des mesures de gestion appropriées à condition d’avoir une compréhension du fonctionnement du système à gérer. Notre premier objectif est donc de mieux mesurer le rôle des relations interspécifiques dans le fonctionnement d’une population, notamment leur interaction avec les traits d’histoire de vie et les conditions environnementales (voir la partie Relations Interspécifiques). Pour les pathogènes par exemple, il est nécessaire de mesurer leur effet sur les paramètres démographiques une fois pris en compte les autres facteurs qui affectent ces paramètres, et de mieux estimer les risques de transmission. Le second objectif est d’analyser les effets des différentes options de gestion, afin de proposer des mesures visant à limiter la propagation de parasites ou le déséquilibre des relations interspécifiques. Ces deux objectifs impliquent une intégration de plus en plus forte entre l’étude des relations interspécifiques et la biologie des populations, notamment pour le développement de modèles complexes.
Évolution de la résistance aux insecticides chez les insectes
Pendant la période qui commence nous comptons approfondir l’étude de l’évolution de la résistance aux insecticides chez les insectes en combinant les développements théoriques avec le travail expérimental, en collaboration avec des collègues de notre unité et extérieurs.
Comme indiqué précédemment, les balanins sont d’importants ravageurs de fruits durs comme les châtaignes, les noisettes, les glands et les noix de pécan. La lutte insecticide contre ces insectes est parfois utilisée mais ne satisfait pas entièrement les exploitants qui sont de plus en plus désireux de produire des fruits « biologiquement » corrects. Ainsi, nous débutons (collaboration F. Menu, S. Venner) un programme de lutte biologique en partenariat avec des structures appliquées comme l’ULRAC (A. Boutitie), un exploitant castanéicole (Mr. Julien, Cévennes) et un producteur de noisette (Mr Ravel, Lyon). Cette lutte nécessite d’intégrer les phénomènes de diapause prolongée des larves et l’existence d’une diapause chez les adultes de certaines espèces. Cette étude appliquée sera donc fortement connectée à nos programmes fondamentaux sur les balanins. Nous proposons de tester l’efficacité de deux types d’auxiliaires : des champignons entomopathogènes et des nématodes.
Perspectives du département
Les perspectives du département pour la période qui commence s’appuient sur les solides compétences développées par les différentes équipes et soulignent la cohérence d’approche de ces équipes au sein du département. Pour chaque sujet, les hypothèses sont clairement identifiées et basées sur des développements théoriques. Nos projets concernent la compréhension de la signification évolutive de différents processus écologiques, le fonctionnement des populations et des systèmes pluri-spécifiques, sans négliger les aspects appliqués de nos travaux qui constituent des réponses aux demandes sociétales (lutte biologique, gestion et conservation). La contribution au développement d’outils méthodologiques ou leur adaptation à de nouvelles problématiques sont et seront aussi une partie importante de nos activités (cf axes transversaux). En tant que département, nos objectifs sont de profiter de la cohérence de nos approches et points de vue pour développer les collaborations entre équipes et avec des collègues étrangers. Ainsi nous proposerons d’organiser, probablement en 2007, un colloque international sur l’écologie évolutive des populations. Une partie du colloque sera dédiée aux implications des aspects évolutifs pour la gestion des populations.
Publications du département (*=avant intégration au laboratoire
2002
P1. Artois M., Manvell R., Fromont E., and Schweyer J.B. (2002) Serosurvey for Newcastle disease and avian influenza A virus antibodies in great cormorants from France. Journal of Wildlife Diseases 38: 169-171.
P2. Bernstein C., Heizmann A., and Desouhant E. (2002) Intraspecific competition between healthy and parasitized hosts in a host parasitoid system : consequences for life-history traits. Ecological Entomology 27: 415-423.
P3. Bonenfant C., Gaillard J.-M., Klein F., and Loison A. (2002) Sex- and age-dependent effects of population density on life history traits of Red deer (Cervus elaphus) in a temperate forest. Ecography 25: 446-458.
P4. Bourdin M., Sallet P., Dufour A.B., and Lacour J.R. (2002) Influence of changes in nasal ventilation on workload during submaximal field running. Journal of Sports Medicine and Physical Fitness 42: 495-499.
P5. Bourgarel M., Fritz H., Gaillard J.-M., Garine-Wichatitsky M.D., and Maudet F. (2002) Effects of annual rainfall and habitat types on the body mass of Impala (Aepyceros melampus) in the Zambezi Valley, Zimbabwe. African Journal of Ecology 40: 186-193.
P6. Bret C., Rahmani A., Dufour A.B., Messonnier L., and Lacour J.R. (2002) Leg strength and stiffness as ability factors in 100-m sprint running. Journal of Sports Medicine and Physical Fitness 42: 274-281.
P7. Calenge C., Maillard D., Gaillard J.-M., Jacobs T., and Merlot L. (2002) Elephant damage to trees of wooded savannah in Zakouma National Park, Chad. Journal of Tropical Ecology 18: 599-614.
P8. Champely S. and Chessel D. (2002) Measuring biological diversity using Euclidean metrics. Environmental and Ecological Statistics 9: 167-177.
P9. Crampe J.P., Gaillard J.-M., and Loison A. (2002) L’enneigement hivernal : un facteur de variation du recrutement chez l’isard (Rupicapra p. pyrenaica). Canadian Journal of Zoology 80: 1306-1312.
P10. Da Silva A. and Allainé D. (2002). Effect of phenotypic and genotypic quality on juvenile alpine marmots survival : preliminary results. in IV Int. Symp. on Alpine Marmot and gen. Marmota. Montreux, Switzerland: 2003.
P11. Debouzie D., Desouhant E., Oberli F., and Menu F. (2002) Resource limitation in natural populations of phytophagous insects. A long-term study case with the chestnut weevil. Acta Oecologica - International Journal of Ecology 23: 31-39.
*P12. Doligez B., Clobert J., Pettifor R.A., Rowcliffe M., Gustafsson L., Perrins C.M., and McCleery R.H. (2002) Costs of reproduction: assessing responses to brood size manipulation on life-history and behavioural traits using multi-state capture-recapture models. Journal of Applied Statistics 29: 407-423.
*P13. Doligez B., Danchin E., and Clobert J. (2002) Public information and breeding habitat selection in a wild bird population. Science 297: 1168-1170.
P14. Dray S., Pettorelli N., and Chessel D. (2002) Multivariate analysis of incomplete mapped data. Transactions in GIS 7: 411-422.
P15. Dray S., Pettorelli N., and Chessel D. (2002) Matching data sets from two different spatial samples. Journal of Vegetation Science 13: 867-874.
P16. Dumonteil E., Gourbière S., Barrera-Perez M., Rodriguez-Félix E., Ruiz-Pina H., Banos-Lopez O., Ramirez-Siera M.J., Menu F., and Rabinovich J.E. (2002) Geographic distribution of Triatoma dimidiata and transmission dynamics of Trypanosoma cruzi in the Yucatan peninsula, Mexico. American Journal of Tropical Medicine and Hygiene 67: 176-183.
P17. Farand E., Allainé D., and Coulon J. (2002) Variation in survival rates of the monogamous alpine marmot (Marmota marmota) : effects of sex, age, year and climate factors. Canadian Journal of Ecology 80: 342-349.
P18. Festa-Bianchet M., Blanchard P., Gaillard J.-M., and Hewison A.J.M. (2002) Tooth extraction is not an acceptable technique to age live ungulates. Wildlife Society Bulletin 30: 282-283.
P19. Hewison A.J.M., Gaillard J.-M., Angibault J.M., Van Laere G., and Vincent J.P. (2002) The influence of density on post-weaning winter growth in roe deer Capreolus capreolus fawns. Journal of Zoology 257: 303-309.
P20. Hewison A., Gaillard J.M., Blanchard P., and Festa-Bianchet M. (2002) Maternal age is not a predominant determinant of progeny sex ratio variation in ungulates. Oïkos 98: 334-339.
P21. King W. and Allainé D. (2002) Dominance relationships, male turnover and other factors influencing reproductive success of female alpine marmots (Marmota marmota). Canadian Journal of Zoology 80: 2137-2143.
P22. Leonard Y., Maillard D., Suisse-Guillaud T., Wanner M., and Calenge C. (2002) La stratégie d’adaptation du chevreuil au milieu montagnard : premiers résultats d’une étude menée dans les Alpes du Sud. Faune Sauvage 257.
P23. Loison A., Cuyler L.C., Linnell J.D.C., and Landa A. (2002) Sex, age and tooth wear of harvested caribou in west Greenland, 1995-1998. Wildlife Biology 7: 263-274.
P24. Loison A., Sæther B.E., and Jerstad K. (2002) Disentangling density and climate effect of dipper survival in Norway. Journal of Applied Statistics 29: 289-304.
P25. Loison A., Toïgo C., Appolinaire J., and Michallet J. (2002) Population dynamics and demographic processes in colonizing populations of ungulates: chamois and ibex as case studies. Journal of Zoology 256: 199-205.
P26. Maillard D., Calenge C., Invernia N., and Gaudin J.C. (2002) Home range size and reproduction of female roe deer re-introduced into a Mediterranean habitat. Zeitschrift für Jagdwissenschaft 48: 194-200.
P27. Menu F. and Desouhant E. (2002) Bet-hedging for variability in life cycle duration: bigger and later-emerging chestnut weevils have increased probability of a prolonged diapause. Oecologia 132: 167-174.
P28. Messonnier L., Freund H., Denis C., Dormois D., Dufour A.B., and Lacour J.R. (2002) Time to exhaustion is related to lactate exchange and removal abilities. International Journal of Sports Medicine 23: 433-438.
P29. Meunier J. and Bernstein C. (2002) The influence of local mate competition on host-parasitoid dynamics. Ecological Modelling 152: 77-88.
P30. Michel J.F., Dray S., de La Rocque S., Desquesnes M., Solano P., De Wispelaere G., and Cuisance D. (2002) Modelling bovine trypanosomosis spatial distribution by GIS in an agro-pastoral zone of Burkina Faso. Preventive Veterinary Medicine 56: 5-18.
P31. Pélissier R., Dray S., and Sabatier D. (2002) Within-plot relationships between tree species occurrences and hydrological soil constraints : an example in French Guiana investigated through canonical correlation analysis. Plant Ecology 162: 143-156.
P32. Pettorelli N., Gaillard J.-M., Van Laere G., Duncan P., and Maillard D. (2002) Cohort effects and spatial variation in adult mass of roe deer. Proceedings of the Royal Society of London. Series B, Biological sciences 269: 747-753.
P33. Pontier D., Say L., Debias F., Bried J., Thioulouse J., Micol T., and Natoli E. (2002) The diet of feral cats (Felix catus L.) at five sites on the Grande Terre, Kerguelen archipelago. Polar Biology 25: 833-837.
P34. Sauvage F., Penalba C., Boué F., Coudrier D., and Artois M. (2002) Puumala hantavirus infection in humans and in the reservoir host, Ardennes region, France. Emerging Infectious Diseases 8: 1509-1511.
P35. Say L., Devillard S., Natoli E., and Pontier D. (2002) The mating system of feral cats (Felix catus L.) in a sub-Antarctic environment. Polar Biology 25: 838-842.
P36. Say L., Gaillard J.-M., and Pontier D. (2002) Spatio-temporal variation in cat population density in a sub-antarctic environment. Polar Biology 25: 90-95.
P37. Schneider M.V., Beukeboom L.W., Driessen G., Lapchin L., Bernstein C., and van Alphen J.J.M. (2002) Geographical distribution and genetic relatedness of sympatrical thelytokous and arrhenotokous populations of the parasitoid Venturia canescens (Hymenoptera). Journal of Evolutionary Biology 15: 191-200.
P38. Skonhoft A., Yoccoz N.G., Stenseth N.C., Gaillard J.-M., and Loison A. (2002) Management of chamois (Rupicapra rupicapra) moving between a protected core area and a hunting area. Ecological Applications 12: 1199-1211.
P39. Solberg E.J., Loison A., Ringsby T.H., Sæther B.E., and Heim M. (2002) Biased adult sex ratio can affect fecundity in primiparous moose Alces alces. Wildlife Biology 8: 117-128.
P40. Toïgo C., Gaillard J.-M., Gauthier D., Girard I., Martinot J.P., and Michallet J. (2002) Female reproductive success and costs in a temperate capital breeder : the effect of contrasting environmental conditions. Ecoscience 9: 427-433.
P41. Valière N., Berthier P., Mouchiroud D., and Pontier D. (2002) Geminy : software for testing the effects of genotyping errors and multitubes approach for the individual identification. Molecular Ecology Notes 2: 83-86.
2003
P42. Allainé D. and Yoccoz, N.G. (2003) Rodents in the European Alps : population ecology and potential impacts on ecosystems, in Alpine Biodiversity in Europe, Nagy L., Grabherr G., Körner C., and Thompson D.B.A., Editors. Springer: Berlin. p.339-349.
P43. Amat I., Bernstein C., and van Alphen J.J.M. (2003) Does a deletion in a virus-like particle protein pleiotropic have effects on the reproductive biology of a parasitoid wasp? Journal of Insect Physiology 49: 1183-1188.
P44. Artois M., Fromont E., and Hars J. (2003) La faune sauvage, indicateur possible du risque de maladie émergente. Epidémiologie et Santé Animale 44: 21-31.
P45. Bahi-Jaber N., Langlais M., and Pontier D. (2003) Behavioral plasticity and virus propagation: the FIV-cat population example. Theoretical Population Biology 64: 11-24.
P46. Bahi-Jaber N. and Pontier D. (2003) Modeling transmission of directly transmitted infectious diseases using colored stochastic Petri nets. Mathematical Biosciences 185: 1-13.
P47. Blanchard P., Festa-Bianchet M., Gaillard J.-M., and Jorgenson J.T. (2003) A test of long-term fecal nitrogen monitoring to evaluate nutritional status in bighorn sheep. Journal of Wildlife Management 67: 477-484.
P48. Bonenfant C., Gaillard J.-M., Loison A., and Klein F. (2003) Sex-ratio variation and reproductive costs in relation to density in a forest dwelling population of red deer (Cervus elaphus). Behavioral Ecology 14: 446-458.
P49. Bret C., Messonnier L., Nouck Nouck J.M., Freund H., Dufour A.B., and Lacour J.R. (2003) Differences in Lactate exchange and removal abilities in athletes specialised in different Track Running Events (100m to 1500m). International Journal of Sports Medicine 24: 108-113.
P50. Bried J., Pontier D., and Jouventin P. (2003) Mate fidelity in monogamous birds : a re-examination of the Procellariiformes. Animal Behaviour 65: 235-246.
P51. Calenge C., Birkan M., and Lebreton P. (2003) Separation of ecological niches of galliform mountain birds in the Northern Alps (Vanoise National Park). Game and Wildlife Science 20: 259-285.
P52. Calenge C., Maillard D., Vassant J., and Brandt S. (2003) Summer and hunting season home ranges of wild boar (Sus scrofa) in two habitats in France. Game and Wildlife Science 19: 281-301.
P53. Casas J., Driessen G., Mandon N., Wielaard B., Desouhant E., Van Alphen J.J.M., Lapchin L., Rivero A., Christides J.P., and Bernstein C. (2003) Energy dynamics in a parasitoid foraging in the wild. Journal of Animal Ecology 72: 691-697.
P54. Castelo M., Corley J.C., and Desouhant E. (2003) Conspecific avoidance during foraging in Venturia canescens (Hymenoptera: Ichneumonidae) : the roles of host presence and conspecific densities. Journal of Insect Behavior 16: 307-318.
P55. Da Silva A., Luikart G., Allainé D., Gautier P., Taberlet P., and Pompanon F. (2003) Isolation and caractérisation of microsatellites in European alpine marmots (Marmota marmota). Molecular Ecology Notes 3: 189-190.
P56. Dechesne A., Pallud C., Debouzie D., Flandrois J.P., Gaudet J.P., and Grundmann G.L. (2003) A method to study the microscale 3-D spatial distribution of Bacteria in soil. Mycological Research 107: 1221-1230.
P57. Dechesne A., Pallud C., Debouzie D., Flandrois J.P., Vogel T.M., Gaudet J.P., and Grundmann G.L. (2003) A novel method for characterizing the microscale spatial distribution of bacteria in soil. Soil Biology and Biochemistry 35: 1537-1546.
P58. Desouhant E., Driessen G., Wielaard B., Lapchin L., and Bernstein C. (2003) Dispersal between host populations in field conditions : navigation rules in the parasitoid Venturia canescens. Ecological Entomology 28: 257-267.
P59. Devillard S., Say L., and Pontier D. (2003) Dispersal pattern of domestic cats (Felix catus) in a promiscuous urban population: do females disperse or die ? Journal of Animal Ecology 72: 203-211.
*P60. Doligez B., Cadet C., Danchin E., and Boulinier T. (2003) When to use public information for breeding habitat selection? The role of environmental predictability and density dependence. Animal Behaviour 66: 973-988.
*P61. Doligez B. and Clobert J. (2003) Experimental increase in nest predation rate induces clutch size reduction in the collared flycatcher. Ecology 84: 2582-2588.
P62. Dray S., Chessel D., and Thioulouse J. (2003) Procustean co-inertia analysis for the linking of multivariate datasets. Ecoscience 10: 110-119.
P63. Dray S., Chessel D., and Thioulouse J. (2003) Co-inertia analysis and the linking of ecological tables. Ecology 84: 3078-3089.
P64. Festa-Bianchet M., Gaillard J.-M., and Côté S.D. (2003) Variable age structure and apparent density-dependence in survival of adult ungulates. Journal of Animal Ecology 72: 640-645.
P65. Fromont E., Pontier D., and Langlais M. (2003) Disease propagation in connected host populations with density-dependent dynamics : the case of the Feline Leukemia Virus. Journal of Theoretical Biology 223: 465-475.
P66. Gaillard J.-M., Duncan P., Delorme D., Van Laere G., Pettorelli N., Maillard D., and Renaud G. (2003) Effects of Hurricane Lothar on the population dynamics of European roe deer. Journal of Wildlife Management 67: 767-773.
P67. Gaillard J.-M., Loison A., Festa-Bianchet M., Yoccoz N.G., and Solberg E.J. (2003b) Ecological Correlates of Life Span in Populations of Large Herbivorous Mammals. Population and Development Review 29: 39-56.
P68. Gaillard J.-M., Loison A., Festa-Bianchet M., Yoccoz N.G., and Solberg E.J. (2003) Ecological correlates of life span in populations of large herbivorous mammals, in Life span : evolutionary and demographic perspectives, Carey J.R., Tuljapurkar S., Editors. Population council: New York. p.39-56.
P69. Gaillard J.-M., Loison A., and Toïgo C. (2003a) Variation in life history traits and realistic population models for wildlife management: the case of ungulates, in Animal behavior and wildlife conservation, Festa-Bianchet M., Appolonio M., Editors. Island press: Washington. p.115-132.
P70. Gaillard J.-M., Loison A., Toïgo C., Delorme D., and Van Laere G. (2003c) Cohort effects and deer population dynamics. Ecoscience 10: 312-320.
P71. Gaillard J.-M. and Yoccoz N.G. (2003) Temporal variation in survival of mammals: a case of environmental canalization? Ecology 84: 3294-3306.
P72. Gimaret-Carpentier C., Dray S., and Pascal J.-P. (2003) Broad-scale biodiversity pattern of the endemic tree flora of the Western Ghats (India) using canonical correlation analysis of herbarium records. Ecography 26: 429-444.
P73. Gourbière F. and Debouzie D. (2003) local variations in microfungal populations on Pinus sylvestris needles. Mycological Research 107: 1221-1230.
P74. Lobry J.R. and Chessel D. (2003) Internal correpondence analysis of codon and amino acid usage in thermophilic bacteria. Journal of Applied Genetics 44: 235-261.
P75. Loison A., Toïgo C., and Gaillard J.-M. (2003) Large herbivore biodiversity in European mountain ecosystems: current status and challenge for the future, in Alpine biodiversity- Pattern, process and change in Europe, Nagy L., Editor. Springer Verlag: Berlin. p. 351-366.
P76. Ollier S., Chessel D., Couteron P., Pelissier R., and Thioulouse J. (2003) Comparing and classifying one-dimensional spatial patterns: an application to laser altimeter profiles. Remote Sensing of Environment 85: 453-462.
*P77. Pärt T. and Doligez B. (2003) Public information and habitat selection: prospecting birds cue on parental activity. Proceedings of the Royal Society of London. Series B, Biological sciences 270: 1809-1813.
P78. Pélissier R., Couteron P., Dray S., and Sabatier D. (2003) Consistency between ordination techniques and diversity measurements: two alternative strategies for species occurrence data. Ecology 84: 242-251.
P79. Pettorelli N., Dray S., Gaillard J.-M., Chessel D., Duncan P., Illius A., Guillon N., Klein F., and Van Laere G. (2003b) Spatial variation in springtime food resources influences the winter body mass of roe deer fawns. Oecologia 137: 363-369.
P80. Pettorelli N., Gaillard J.-M., Duncan P., Maillard D., Van Laere G., and Delorme D. (2003a) Age and density modify the effects of habitat quality on survival and movements of roe deer. Ecology 84: 3307-3316.
P81. Poussin J.C., Wopereis M.C.S., Debouzie D., and Maeght J.L. (2003) Determinants of irrigated rice yield in the Senegal river. European Journal of Agronomy 19: 341-356.
P82. Sauvage F., Langlais M., Yoccoz N.G., and Pontier D. (2003) Modelling hantavirus in fluctuating populations of bank voles: the role of indirect transmission on virus persistence. Journal of Animal Ecology 72: 1-13.
P83. Say L., Bonhomme F., Desmarais E., and Pontier D. (2003) Microspatial genetic heterogeneity and gene flow in stray cats (Felix catus L.): a comparison of coat colour and microsatellite loci. Molecular Ecology 12: 1669-1674.
P84. Say L., Naulty F., and Hayden T.J. (2003) Genetic and behavioural estimates of reproductive skew in male fallow deer. Mol. Ecol. 12 : 2793-2800.
P85. Soula B. and Menu F. (2003) Variability in diapause duration in the chestnut weevil: mixed ESS, genetic polymorphism or bet-hedging ? Oïkos 100: 574-580.
P86. Thioulouse J., Simier M., and Chessel D. (2003) Simultaneous analysis of a sequence of pairs of ecological tables with the STATICO method. Ecology 85: 272-283.
P87. Toïgo C. and Gaillard J.-M. (2003) Causes of sex-biased adult survival in ungulates : sexual size dimorphism, mating tactic or environment harshness ? Oïkos 101: 376-384.
P88. van Alphen J.J.M., Bernstein C., and Driessen G. (2003) Information acquisition and time allocation in insect parasitoids. Trends in Ecology and Evolution 18: 81-87.
P89. Venner S., Bel-Venner M.C., Pasquet A., and Leborgne R. (2003) Body mass-dependent cost of web-building behaviour in an orb weaving spider Zygiella x-notata. Naturwissenschaften 90: 269-272.
2004
P90. Allainé D. (2004) Sex ratio variation in the cooperatively breeding alpine marmot Marmota marmota. Behavioral Ecology 15: 987-1002.
P91. Allainé D. and Theuriau F. (2004) Is there an optimal number of helpers in alpine marmot family groups? Behavioral Ecology 15: 916-924.
P92. Bonenfant C., Gaillard J.-M., Klein F., and Maillard D. (2004) Variation in harem size of red deer (Cervus elaphus) : the effects of adult sex ratio and age structure. Journal of Zoology 264: 1-9.
P93. Bonenfant C., Loe L.E., Mysterud A., Langvatn R., Stenseth N.C., Gaillard J.-M., and Klein F. (2004) Multiple causes of sexual segregation in European red deer : enlightenments from varying breeding phenology at high and low latitude. Proceedings of the Royal Society of London. Series B, Biological sciences 271: 883-892.
P94. Calenge C. (2004) Efficiency of spreading maize in the garrigues to reduce wild boar damage to Mediterranean vineyard. European Journal of Wildlife Research 50: 112-120.
P95. Chessel D., Dufour A.-B., and Thioulouse J. (2004) The ade4 package-I- One-table methods. R News 4: 5-10.
P96. Corley J.C., Capurro A.F., and Bernstein C. (2004) Prolonged diapause and the stability of host-parasitoid interactions. Theoretical Population Biology 65: 193-203.
P97. Devillard S., Allainé D., Gaillard J.-M., and Pontier D. (2004a) Does social complexity lead to sex-biased dispersal in polygynous mammals ? A test on ground-dwelling sciurids. Behavioral Ecology 15: 83-87.
P98. Devillard S., Say L., and Pontier D. (2004b) Molecular and behavioural analyses reveal male-biased dispersal between social groups of domestic cats. Ecoscience 11: 175-180.
P99. Doligez B., Pärt T., and Danchin E. (2004) Prospecting in the collared flycatcher: gathering public information for breeding habitat selection? Animal Behaviour 67: 457-466.
P100. Doligez B., Pärt T., Danchin E., Clobert J., and Gustafsson L. (2004) Availability and use of public information and conspecific density for settlement decisions in the collared flycatcher. Journal of Animal Ecology 73: 75-87.
P101. Doligez B., Thomson D.L., and van Noordwijk A.J. (2004) Using large-scale data analysis to assess life history and behavioural traitrs: the case of the reintroduced White stork population in the Netherlands. Animal Biodiversity and Conservation 27: 387-402.
P102. Dumonteil E., Ruiz-Pina H., Rodriguez-Félix E., Barrera-Perez M., Ramirez-Sierra M.J., Rabinovich J.E., and Menu F. (2004) Re-infestation of houses by Triatoma dimidiata after intra-domicile insecticide application in the Yucatan Peninsula, Mexico. Mem. Inst. Oswaldo Cruz, Rio de Janeiro 99: 253-256.
P103. Faurie C., Pontier D., and Raymond M. (2004) Student athletes claim to have more sexual partners than other students. Evolution and Human Behavior 25: 1-8.
P104. Fitzgibbon, W.E., Langlais M., Morgan J.J., Pontier D., and Wolf C. (2004) An age-dependent model describing the spread of panleucopenia virus within feline populations. Banach Center Publications, Polish Academy of Science 63: 197-207.
P105. Forsyth D.M., Tustin, K.G., Gaillard J.-M., and Loison A. (2004) Fetal sex ratio variation in the highly polygynous Himalayan tahr : evidence for differential mortality. Behavioral Ecology 15: 572-578.
P106. Fournier-Chambrillon C., Bent A., Pérrot A., Pontier D., Sauvage F., Artois M., and Fournier P. (2004) Occurrence of antibodies to Aleutian Mink Disease Parvovirus in free-ranging small carnivores from south-western France : implication for the conservation of the European mink (Mustela lutreola). Journal of Wildlife Diseases 40: 394-402.
P107. Garel M., Loison A., Gaillard J.-M., Cugnasse J.-M., and Maillard D. (2004) The effects of a severe drought on mouflon lamb survival. Proceedings of the Royal Society of London. Series B, Biological sciences 271: S471-S473.
P108. Grange S., Duncan P., Gaillard J.-M., Sinclair A.R.E., Gogan P.J.P., Packer C., Hofer H., and East M. (2004) What limits the Serengeti zebra population ? Oecologia 140: 523-532.
P109. Halkett F., Harrington R., Hulle M., Kindlmann P., Menu F., Rispe C., and Plantegenest M. (2004) Dynamics of production of sexual forms in Aphids: theoretical and experimental evidence for adaptive "Coin-flipping" plasticity. The American Naturalist 163: E112-E125.
P110. Kaeuffer R., Pontier D., Devillard S., and Perrin N. (2004) Effective size of two feral domestic cat populations (Felix catus L.) : effect of the mating system. Molecular Ecology 13: 483-490.
P111. Kjellander P., Gaillard J.-M., Hewison A.J.M., and Liberg, O. (2004) Predation risk and longevity influence variation in fitness of female roe deer (Capreolus capreolus L.). Proceedings of the Royal Society of London. Series B, Biological sciences 271: 338-340.
P112. Lett C., Auger P., and Gaillard J.-M. (2004) Continuous cycling of grouped vs solitary strategy frequencies in a predator-prey model. Theoretical Population Biology 65: 263-270.
P113. Loison A., Solberg E.J., Yoccoz N.G., and Langvatn R. (2004) Interplay between mother quality and cohort effects on red deer calves. Ecology 85: 1992-2002.
P114. Loison A. and Strand O. (2004) Allometry and variability of resource allocation to reproduction in a wild reindeer population. Behavioral Ecology 27: 47-58.
P115. Molinari-Jobin A., Molinari P., Loison A., Gaillard J.-M., and Breitenmoser U. (2004) Life cycle period and activity of prey influence their susceptibility to predators. Ecography 27: 323-329.
P116. Pallud C., Dechesne A., Gaudet J.P., Debouzie D., and Grundmann G.L. (2004) Modification of spatial distribution of 2,4-dichlorophenooxyacetic acid degrader microhabitats during growth in soil columns. Applied Environment Microbiology 70: 2709-2716.
P117. Pavoine S., Dufour A.-B., and Chessel D. (2004) From dissimilarities among species to dissimilarities among communities : A Double Principal Coordinate Analysis. Journal of Theoretical Biology 228: 523-537.
P118. Raymond M. and Pontier D. (2004) Is there geographical variation in human handedness ? Laterality 9: 35-52.
P119. Say L. and Pontier D. (2004) Spacing pattern in a social group of stray cats : effects on male reproductive success. Animal Behaviour 68: 175-180.
P120. Solberg E.J., Loison A., Gaillard J.-M., and Heim M. (2004) Lasting effects of conditions at birth on moose body mass. Ecography 27: 677-687.
P121. Spataro T. and Bernstein C. (2004) Combined effects of intraspecific competition and parasitoid attacks on the dynamics of a host population: a stage-structured model. Oïkos 105: 148-158.
P122. Spichiger R., Calenge C., and Bise B. (2004) Geographical zonation in the Neotropics of tree species characteristic of the Paraguay-Paraná Basin. Journal of Biogeography 31: 1489 - 1501.
P123. Widmer O., Saïd S., Miroir J., Duncan P., Gaillard J.-M., and Klein F. (2004) The effects of hurricane Lothar on habitat use of roe deer. Forest Ecology and Management 195: 237-242.
2005
P124. Amat I., Castelo M., Desouhant E., and Bernstein C. (2005) The influence of the temperature and host availability on the host exploitation strategies of sexual and asexual parasitic wasps of the same species. Oecologia. Sous presse.
P125. Bel-Venner M.C. and Venner S. (2005) Mate-guarding strategies and male competitive ability : results from a field study in an orb-weaving spider. Animal Behaviour. in press.
P126. Blanchard P. (2005) On lactation and rumination in bighorn ewes (Ovis canadensis). Journal of Zoology 265: 107-112.
P127. Blanchard P., Festa-Bianchet M., Gaillard J.-M., and Jorgenson J.T. (2005) Maternal condition and offspring sex ratio in polygynous ungulates : a case study of bighorn sheep. Behavioral Ecology 16: 274-279.
P128. Bonenfant C., Gaillard J.-M., Klein F., and Hamann J.L. (2005) Can we use the young : female ratio to infer ungulate population dynamics ? An empirical test using red deer (Cervus elaphus) as a model. Journal of Applied Ecology 42: 361-370.
P129. Brenot J.F., Ellison L., Rotelli L., Novoa C., Calenge C., Léonard P., and Menoni E. (2005) Is body mass of rock ptarmigan greater in the Pyrenees than in the Alps ? Wildlife Biology. Sous Presse.
P130. Calenge C., Dufour A.-B., and Maillard D. (2005) K-select analysis : a new method to analyse habitat selection in radio-tracking studies. Ecological Modelling 186: 143-153.
P131. Calenge C., Maillard D., Invernia N., and Gaudin J.C. (2005) Reintroduction of roe deer into a Mediterranean habitat : female mortality and dispersion. Wildlife Biology. Sous presse.
P132. Cohas A., Yoccoz N.G., Da Silva A., Goossens B., and Allainé D. (2005) Extra-pair paternity in the monogamous alpine marmot (Marmota marmota) : The roles of social setting and female mate choice. Behavioral Ecology and Sociobiology. sous presse.
P133. Coulson T., Gaillard J.-M., and Festa-Bianchet M. (2005) Decomposing the variation in population growth into contributions from multiple vital rates. Application to bighorn sheep and red deer. Journal of Animal Ecology 74: 789-801.
P134. Couteron P. and Ollier S. (2005) A generalized, variogram-based framework for multi-scale ordination. Ecology 86: 828-834.
P135. Da Silva A., Luikart G., Yoccoz N.G., Cohas A., and Allainé D. (2005) Genetic diversity-fitness correlation revealed by microsatellite analyses in European alpine marmots (Marmota marmota). Conservation Genetics. in press.
P136. De Palma C., Viggiano E., Barillari E., Palme R., Dufour A.B., Fantini C., and Natoli E. (2005) Evaluating the temperament in shelter dogs. Journal of Behavior.
P137. Desouhant E., Driessen G., Amat I., and Bernstein C. (2005) Host and food searching in a parasitic wasp Venturia canescens : a trade-off between current and future reproduction ? Animal Behaviour 70: 145-152.
P138. Forsyth D.M., Duncan R.P., Tustin K.G., and Gaillard J.-M. (2005) A substantial energetic cost to male reproduction in a sexually dimorphic ungulate. Ecology 86: 2154-2163.
P139. Gaillard J.-M. (2005) Is moose just a large deer species ? Some life history considerations. Alces. Sous presse.
P140. Gaillard J.-M., Viallefont A., Loison A., and Festa-Bianchet M. (2005) Assessing senescence patterns in populations of large mammals. Animal Biodiversity and Conservation 27: 47-58.
P141. Gaillard J.-M., Yoccoz N.G., Lebreton J.-D., Bonenfant C., Devillard S., Loison A., Pontier D., and Allainé D. (2005) Generation time : a reliable metric to measure life history variation among mammalian populations. American Naturalist 166:: 119-123.
P142. Garel M., Cugnasse J.-M., Gaillard J.-M., Loison A., Gibert P., Douvre P., and Dubray D. (2005) Reproductive output of female mouflon : a comparative analysis. Journal of Zoology 266: 65-71.
P143. Garel M., Cugnasse J.-M., Gaillard J.-M., Loison A., Santosa Y., and Maublanc M.L. (2005) Effects of observer experience on the monitoring of a Mediterranean mouflon population. Acta Theriologica 50: 109-114.
P144. Garel M., Cugnasse J.-M., Loison A., Gaillard J.-M., Vuiton C., and Maillard D. (2005) Monitoring the abundance of mouflon in South France. European Journal of Wildlife Research 51: 69-76.
P145. Gaucel S., Langlais M., and Pontier D. (2005) Invading introduced species in insular heterogeneous environments. Ecological Modelling. in press.
P146. Gaucel S. and Pontier D. (2005) How predator food preference can change the destinity of native prey in predator-prey systems. Biological Invasions. in press.
P147. Haguenauer M., Fargier P., Legreneur P., Dufour A.B., Cogerino G., Begon M., and Monteil K.M. (2005) Short-term effects of using verbal instructions and demonstration at the beginning of learning a complex skill figure skating. Perceptual and Motor Skills 100 : 179-91.
P148. Henriksen H.B., Andersen R., Hewison A.J.M., Gaillard J.-M., Bronndal M., Jonsson S., Linnell J.D.C., and Odden J. (2005) Reproductive biology of captive female Eurasian lynx, Lynx lynx. European Journal of Wildlife Research 51: 151-156.
P149. Hewison A.J.M., Gaillard J.-M., Kjellander P., Toïgo C., Liberg O., and Delorme D. (2005) Big mothers invest more in daughters - reversed sex allocation in a weakly polygynous mammal. Ecology Letters 8: 430-437.
P150. Kjellander P., Gaillard J.-M., and Hewison A.J.M. (2005) Density-dependent responses of fawn cohort body mass in two contrasting roe deer populations ? Oecologia. in press.
P151. Loe L.E., Bonenfant C., Mysterud A., Gaillard J.-M., Langvatn R., Stenseth N.C., Klein F., and Pettorelli N. (2005) Climate predictability and breeding phenology in red deer : timing and synchrony of rutting and calving in Norway and France. Journal of Animal Ecology 74: 579-588.
P152. Loison A., Appolinaire J., Jullien J.M., and Dubray D. (2005) How reliable are population counts to detect trends in population size of chamois ? Wildlife Biology. in press.
P153. Natoli E., Say L., Cafazzo S., Bonanni R., Schmid M., and Pontier D. (2005) Bold attitude makes male urban feral domestic cats more vulnerable to Feline Immunodeficiency Virus. Neuroscience and Biobehavioral Reviews 29: 151-157.
P154. Ney-Nifle M., Bernstein C., Reboreda J.C., and Kacelnik A. (2005) Population dynamics and avian brood parasitism : persistence and invasions in a three species system. Journal of Animal Ecology 74: 274-284.
P155. Ollier S., Couteron P., and Chessel D. (2005) Orthonormal transform to detect and characterize phylogenetic signal. Biometrics. in press.
P156. Pavoine S. and Dolédec S. (2005) The apportionment of quadratic entropy: a useful alternative for partitioning diversity in ecological data. Environmental and Ecological Statistics 12: 125-138.
P157. Pavoine S., Ollier S., and Dufour A.-B. (2005) Is the originality of a species measurable ? Ecology Letters 8: 579-586.
P158. Pavoine S., Ollier S., and Pontier D. (2005) Measuring diversity from dissimilarities with Rao’s quadratic entropy : are any dissimilarity indices suitable ? Theoretical Population Biology 67: 231-239.
P159. Pettorelli N., Gaillard J.-M., Mysterud A., Duncan P., Stenseth N.C., Delorme D., Van Laere G., Toïgo C., and Klein F. (2005a) Using a proxy of plant productivity (NDVI) to find key periods for animal performance : the case of roe deer. Oïkos. in press.
P160. Pettorelli N., Gaillard J.-M., Yoccoz N.G., Duncan P., Maillard D., Delorme D., Van Laere G., and Toïgo C. (2005b) The response of fawn survival to changes in habitat quality varies according to cohort quality and spatial scale. Journal of Animal Ecology 74: 972-981.
P161. Pettorelli N., Vik J.O., Mysterud A., Gaillard J.-M., Tucker C.J., and Stenseth N.C. (2005c) Using the satellite-derived Normalized Difference Vegetation Index (NDVI) to assess responses of animal populations to environmental change. Trends in Ecology and Evolution. in press.
P162. Plenet S., Joly P., Hervant E., Fromont E., and Grolet O. (2005) Are hybridogenetic zones structured by environmental gradients? Evidence from in situ experiments in the waterfrog hybridization complex. Journal of Evolutionary Biology. sous presse.
P163. Pontier D., Say L., and Bonhomme F. (2005) Population genetic structure of the feral cat Felis catus L. on a subantarctic Island 50 years after introduction. Polar Biology 28: 268-275.
P164. Richomme C., Fromont E., and Gauthier D. (2005) Contact rates and exposure to interspecies disease transmission in mountain ungulates. Epidemiology and Infection. in press.
P165. Rossi S., Artois M., Pontier D., Crucière C., Hars J., Barrat J., Pacholek X., and Fromont E. (2005a) Long-term monitoring of classical swine fever in wild boar (Sus scrofa sp.) using serological data. Veterinary Research 36: 27-42.
P166. Rossi S., Fromont E., Pontier D., Crucière C., Hars J., Barrat J., Pacholek X., and Artois M. (2005b) Incidence and persistence of classical swine fever in free-ranging wild boar (Sus scrofa). Epidemiology and Infection 133: sous presse.
P167. Saïd S., Pellerin M., Guillon N., Debias F., and Fritz H. (2005) Assessment of forage availability in ecological studies. European Journal of Wildlife Research in press.
P168. Saïd-Delcros S., Gaillard J.-M., Duncan P., Guillon N., Guillon N., Servanty S., Pellerin M., Lefeuvre K., Martin C., and Van Laere G. (2005) Ecological correlates of home-range size in spring-summer for female roe deer (Capreolus capreolus) in a deciduous woodland. Journal of Zoology. In press.
P169. Say L., Naulty F., Fennell A., and Hayden T.J. (2005) Absence of heterozygote advantage in a population of fallow deer. Biological Letters (sous presse).
P170. Soula B. and Menu F. (2005) Extended life cycle in the chestnut weevil : prolonged diapause or repeated diapause ? Entomologia Experimentalis et Applicata 115 : 333-340.
P171. Tentelier C., Desouhant E., and Fauvergue X. (2005) Habitat assessment by parasitoids : mechanisms for patch use behavior. Behavioral Ecology. en révision: 00-00.
P172. Venner S. and Casas J. (2005) Spider webs designed for rare but life-saving catches. Proceedings of the Royal Society of London B 272: 1587-1592.
P173. Zanneïse A., Baïsse A., Gaillard J.-M., Hewison A.J.M., Saint-Hilaire K., Toïgo C., Van Laere G., and Morellet N. (2005) Hind foot length : an indicator for monitoring roe deer populations at a landscape scale. Wildlife Society Bulletin. In press.